海蜇生活史的補(bǔ)充研究
2014-12-15 03:34:58劉春勝莊志猛陳四清劉長(zhǎng)琳
海洋科學(xué) 2014年12期
劉春勝, 莊志猛, 陳四清, 劉長(zhǎng)琳
(中國(guó)水產(chǎn)科學(xué)研究院 黃海水產(chǎn)研究所, 山東 青島 266071)
海蜇(Rhopilema esculentum)隸屬于腔腸動(dòng)物門(Cnidaria)、缽水母綱(Scyphozoa)、根口水母目(Rhizostomeae)、根口水母科(Rhizostomatidae)、海蜇屬(Rhopilema), 分布于中國(guó)、日本、朝鮮沿岸以及俄羅斯遠(yuǎn)東水域[1-3]。作為品質(zhì)最好的食用水母, 海蜇是中國(guó)最重要的海水漁業(yè)增養(yǎng)殖對(duì)象之一, 同時(shí)也是國(guó)際上唯一的人工養(yǎng)殖類水母。中國(guó)是最早開展海蜇研究的國(guó)家, 據(jù)報(bào)道, 早在20世紀(jì)60年代就對(duì)海蜇資源進(jìn)行了深入調(diào)查, 并于80年代首次報(bào)道了海蜇的生活史及開展人工繁育實(shí)驗(yàn)[4]。可以說, 中國(guó)在海蜇研究中始終處于世界的前列。
作為進(jìn)化史上真正的兩胚層動(dòng)物, 缽水母的生活史非常復(fù)雜, 包括附著的螅狀體階段和浮游的水母體階段, 以及兩者交替轉(zhuǎn)變過程中的浮浪幼蟲階段和橫裂體階段[5]。因此, 明晰復(fù)雜生活史各個(gè)階段的形態(tài)特征, 詳細(xì)劃分生活史的典型階段, 對(duì)于開展缽水母的基礎(chǔ)生物學(xué)研究具有重要意義。然而, 在超過200多種已鑒定缽水母中, 僅有17種水母的完整生活史被報(bào)道[6]。導(dǎo)致該現(xiàn)象的主要原因是缽水母的螅狀體及初生浮游幼體階段個(gè)體非常小, 且某些階段歷時(shí)短, 如浮浪幼蟲、碟狀體等階段。此外, 含水量高(超過95%)、形態(tài)極易破損也是導(dǎo)致水母類生活史難以觀察的原因之一。
丁耕蕪等[4]在20世紀(jì)80年代首次報(bào)道了海蜇生活史。然而, 近年來隨著水母在中國(guó)近海的不斷暴發(fā),其在海洋生態(tài)系統(tǒng)中的重要性不斷凸顯, 使得水母旺發(fā)機(jī)制研究成為亟待解決的問題。此后, 中國(guó)近海主要水母物種黃斑海蜇(Rhopilema hispidum)、白色霞水母(Cyanea nozakii)、沙海蜇(Nemopilema nomurai)的生活史被研究報(bào)道[7-10]。在上述水母生活史的相關(guān)研究中, 海蜇總是被作為模式物種進(jìn)行比對(duì)分析。然而, 隨著對(duì)海蜇和其他缽水母種類研究的不斷深入, 現(xiàn)有海蜇生活史各階段的分類已不能滿足某些相關(guān)研究領(lǐng)域的需求, 如在各生活史形態(tài)描述中, 尤其是浮浪幼蟲到幼蟄的幼體階段, 難以對(duì)海蜇與其他缽水母進(jìn)行區(qū)別[7,11]; 在海蜇幼體攝食牙鲆仔稚魚實(shí)驗(yàn)中, 海蜇幼體中央口未閉合時(shí), 能夠攝食牙鲆(Paralichthys olivaceus), 一旦中央口閉合,其對(duì)牙鲆仔稚魚的攝食能力就完全喪失[12]; 劉春勝等[13]在研究浮浪幼蟲攝食時(shí)發(fā)現(xiàn)整個(gè)浮浪幼蟲活力是呈現(xiàn)先升高后下降的趨勢(shì)。綜上, 對(duì)原有海蜇生活史補(bǔ)充完善變得十分迫切和必要, 但是目前尚未有相關(guān)文獻(xiàn)報(bào)道。此外, 作者在近幾年研究中也發(fā)現(xiàn)了許多海蜇特定發(fā)育階段的新現(xiàn)象, 因此本研究將系統(tǒng)觀察海蜇生活史的各個(gè)階段, 并對(duì)其原有生活史進(jìn)行補(bǔ)充、細(xì)化。實(shí)驗(yàn)的開展將使海蜇生活史進(jìn)一步完善, 為開展海蜇基礎(chǔ)生物學(xué)和基礎(chǔ)生態(tài)學(xué)研究提供參考。
1 材料與方法
1.1 實(shí)驗(yàn)材料
2010~2013 年先后在榮成好當(dāng)家水產(chǎn)開發(fā)有限公司、山東省海洋捕撈生產(chǎn)管理站及青島金沙灘水產(chǎn)開發(fā)有限公司開展海蜇有性繁殖實(shí)驗(yàn), 獲取浮浪幼蟲; 實(shí)驗(yàn)室長(zhǎng)期保存螅狀體和足囊; 將低溫保存螅狀體緩慢升溫至 18 ℃, 誘導(dǎo)其橫裂產(chǎn)生碟狀體;實(shí)驗(yàn)室培養(yǎng)獲得各階段海蜇幼體。
1.2 實(shí)驗(yàn)方法
待海蜇親體產(chǎn)卵后, 從池底通過虹吸作用將受精卵轉(zhuǎn)移到2 L玻璃燒杯中, 微充氣, 使受精卵在燒杯中均勻分布。待受精后約6 h, 受精卵發(fā)育至囊胚期開始觀察記錄。每2 h 用顯微鏡觀察1次, 同時(shí)測(cè)量浮浪幼蟲的縱軸長(zhǎng)度及活力。每次觀測(cè)浮浪幼蟲不少于 30個(gè)。浮浪幼蟲活力, 即為自然狀態(tài)下, 浮浪幼蟲每分鐘的旋轉(zhuǎn)圈數(shù)(C/min): 每次隨機(jī)觀察 10個(gè)浮浪幼蟲未受刺激時(shí) 1 min 的旋轉(zhuǎn)次數(shù), 計(jì)算平均數(shù)。
待受精后約 28 h, 浮浪幼蟲開始附著變態(tài)后,每12 h用顯微鏡觀測(cè)1次, 每次觀測(cè)數(shù)不少于30個(gè)。
足囊萌發(fā): 挑選200個(gè)直徑500 μm以上的飽滿的足囊, 在溫度25 ℃±1 ℃、鹽度 22±1、黑暗條件下誘導(dǎo)足囊萌發(fā), 待發(fā)現(xiàn)萌發(fā)足囊后, 用體視鏡跟蹤觀察。實(shí)驗(yàn)共計(jì)觀測(cè)萌發(fā)足囊34個(gè)。
將低溫度保存螅狀體緩慢升溫至 18℃后, 每天投喂適量鹵蟲(Artemia)無節(jié)幼體, 待出現(xiàn)橫裂體后,用體視鏡跟蹤觀察, 共計(jì)觀測(cè)橫裂體超過100個(gè)。
將橫裂產(chǎn)生的碟狀體轉(zhuǎn)移至 200 L的圓形亞克力缸中, 碟狀體養(yǎng)殖密度為 10個(gè)/L, 每天投喂適量鹵蟲無節(jié)幼體, 喂食后2 h換水2/3。實(shí)驗(yàn)開始后, 每3 d觀測(cè)1次, 直至其發(fā)育至20~30 mm的幼蜇。用體視鏡觀察、拍照記錄。
2 結(jié)果
2.1 浮浪幼蟲形態(tài)變化
海蜇受精卵(圖1a)發(fā)育至囊胚后期逐漸上浮,開始緩慢繞縱軸左旋自傳, 同時(shí)沿自旋軸緩慢拉伸,發(fā)育至浮浪幼蟲期。在整個(gè)浮浪幼蟲階段, 其縱軸是不斷拉伸的, 縱軸由起始階段的100 μm ±21 μm發(fā)育為附著前期的220 μm±26 μm; 伴隨著浮浪幼蟲縱軸的不斷拉伸, 其自旋速度也呈現(xiàn)出先加速后下降的趨勢(shì), 結(jié)果表明在發(fā)育至約 10 h時(shí), 浮浪幼蟲的活力達(dá)到最強(qiáng), 自旋速度為21 C/min±3.2 C/min, 一旦受刺激后還可以通過加速旋轉(zhuǎn), 或通過右旋等方式改變運(yùn)動(dòng)方向以逃避敵害(圖2)。基于發(fā)育過程中海蜇浮浪幼蟲的形態(tài)和旋轉(zhuǎn)速度變化, 其明顯分為3個(gè)階段, 即早期浮浪幼蟲階段(0~4 h), 其特征是浮浪幼蟲縱軸稍長(zhǎng)于短軸, 其前后不易區(qū)分, 旋轉(zhuǎn)緩慢, 對(duì)外界刺激反應(yīng)能力差(圖1b); 中期浮浪幼蟲階段(5~14 h), 其特征是縱軸幾乎為短軸的兩倍, 前端明顯比后端寬, 此時(shí)旋轉(zhuǎn)速度最快, 對(duì)外界刺激反應(yīng)十分劇烈(圖1c); 后期浮浪幼蟲階段(15~32 h), 其特征是縱軸為短軸的 2倍以上, 前端明顯比后端寬,旋轉(zhuǎn)速度逐漸降低, 對(duì)外界反應(yīng)較為遲鈍, 此時(shí)一旦接觸適宜附著基便附著變態(tài)(圖1 d)。
2.2 杯狀體階段
浮浪幼蟲附著變態(tài)(圖3 a~圖3d)和足囊萌發(fā)(圖3 e~圖3 h)是海蜇四觸手螅狀體形成的兩種方式。通過對(duì)這兩種螅狀體形成方式的觀察, 實(shí)驗(yàn)發(fā)現(xiàn)兩者存在許多的共同點(diǎn)。如圖3所示, 均形成類似于高腳酒杯狀的杯狀體結(jié)構(gòu), 該階段的特征是附著部位細(xì)長(zhǎng), 頂部呈現(xiàn)球形, 此階段的海蜇是不攝食的, 兩種方式均是通過內(nèi)源性營(yíng)養(yǎng)完成該階段的發(fā)育(圖3 a, 圖3b, 圖3 e); 之后杯狀體進(jìn)一步發(fā)育, 在球形頂部四周出現(xiàn)4個(gè)凸起, 為四條觸手雛形, 作者稱之為觸手原基(圖3 c, 圖3 f)。隨著杯狀體的進(jìn)一步發(fā)育, 逐漸形成四觸手螅狀體(圖3 d, 圖3 g, 圖3 h)。

圖1 海蜇受精卵和各階段浮浪幼蟲Fig.1 Fertilized egg and planulae at different stages
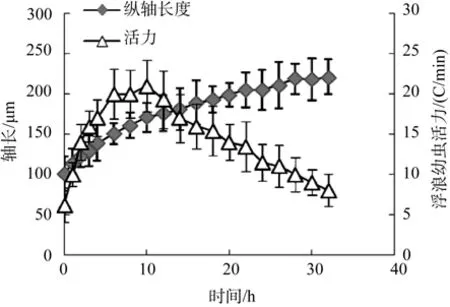
圖2 不同發(fā)育時(shí)期浮浪幼蟲的縱軸長(zhǎng)度和活力Fig.2 Vertical-axis length and vigor of planula at different developmental stages
另外, 海蜇螅狀體兩種發(fā)生方式也存在著明顯的區(qū)別。首先浮浪幼蟲附著變態(tài)形成的螅狀體明顯小于足囊萌發(fā)形成的螅狀體, 因此, 兩種螅狀體的開口餌料也不相同, 前者為約100 μm的海蜇浮浪幼蟲及其他適口餌料, 后者能夠直接捕食約200 μm鹵蟲無節(jié)幼體; 其次兩種方式形成的螅狀體附著點(diǎn)明顯不同, 前者附著點(diǎn)僅為10 μm ~20 μm, 后者足囊營(yíng)養(yǎng)消耗后在其附著點(diǎn)周圍有一圈直徑為原足囊大小的黃色或白色外囊。

圖3 海蜇四觸手螅狀體兩種發(fā)生方式Fig.3 Two generation modes of four tentacles polyp of Rhopilema esculentum
2.3 碟狀體橫裂
即將釋放碟狀體的橫裂體攝食鹵蟲無節(jié)幼體后,能夠直接從最頂端碟狀體穿過, 進(jìn)入到下一個(gè)碟狀體體內(nèi), 因此海蜇橫裂體體腔是上下通透的(圖4a,圖4b); 進(jìn)一步觀察發(fā)現(xiàn)碟狀體橫裂初期也是上下貫通的, 初生碟狀體傘頂部均存在一個(gè)直徑約200 μm的中央洞, 剛好與下一個(gè)碟狀體的口徑一致(圖4c,圖4d), 大約2~3 h后該洞將完全愈合(圖4e, 圖4f)。
2.4 浮游幼體不同階段的劃分
在適宜條件下, 海蜇碟狀體(圖5a)經(jīng)4~5 d后, 以中央洞愈合處為中心, 傘部不斷加厚形成厚的中膠層;同時(shí), 其碟狀體緣瓣間產(chǎn)生行叢輻緣瓣使傘經(jīng)不斷增大, 4對(duì)片狀口腕順中央口下垂, 形成八葉口腕, 海蜇進(jìn)入稚蜇期。在稚蜇期伴隨著八葉口腕的形成, 海蜇中央口開始逐漸封閉, 同時(shí)口腕周圍形成細(xì)小的吸口雛形。實(shí)驗(yàn)根據(jù)中央口和吸口的愈合封閉與否將稚蜇分為稚蜇前期和稚蜇后期。開放的中央口是稚蜇前期的特征之一, 該階段的海蜇可以同時(shí)通過新形成的吸口和中央口攝食, 中央口的直徑可達(dá)1 mm以上, 因此該階段海蜇能夠直接捕食仔稚魚等大規(guī)格浮游生物, 對(duì)我國(guó)沿海某些漁業(yè)資源有直接影響(圖5b, 圖5c)。
隨著海蜇浮游幼體的繼續(xù)發(fā)育, 其中央口完全封閉, 進(jìn)入稚蜇后期, 該階段海蜇只能通過周圍吸口攝食(圖5 d, 圖5e, 圖5f)。
當(dāng)稚蜇生長(zhǎng)到傘徑長(zhǎng)到約 20 mm, 口腕基本愈合, 口腕外端長(zhǎng)出棒狀附著器, 此時(shí)海蜇浮游幼體形態(tài)特征與成體水母基本一致, 這個(gè)階段的幼體稱為幼蜇(圖5 g, 圖5h)。

圖4 碟狀體橫裂Fig.4 Strobilation of ephyra

圖5 各階段海蜇浮游幼體Fig.5 planktonic larvae of R.esculentum at different stages
3 討論與結(jié)論
缽水母生活史均可分為有性繁殖的水母體階段和無性繁殖的螅狀體階段, 整個(gè)生活史過程包括胚胎發(fā)育(浮浪幼蟲階段)、附著變態(tài)(螅狀體階段)、無性繁殖(足囊繁殖、出芽繁殖及橫裂生殖)及浮游水母體階段。但與魚類等有明確生活史分期標(biāo)準(zhǔn)的物種不同, 至今, 缽水母生活史尚未有一定的標(biāo)準(zhǔn)。作為中國(guó)最早被研究的缽水母, 海蜇的生活史于80年代被報(bào)道[4]。本研究在此基礎(chǔ)上, 通過顯微觀察和比較分析, 依據(jù)海蜇各發(fā)育階段的形態(tài)變化, 對(duì)海蜇整個(gè)生活史階段進(jìn)行細(xì)致地劃分, 報(bào)道了其在浮浪幼蟲發(fā)育、四觸手螅狀體形成、碟狀體橫裂及浮游幼體發(fā)育4個(gè)階段中的新現(xiàn)象或新階段。
3.1 浮浪幼蟲不同階段的劃分
有學(xué)者認(rèn)為類似浮浪幼蟲的祖先是兩層動(dòng)物細(xì)胞到兩胚層動(dòng)物細(xì)胞過渡的典型代表[14-15]。因此, 對(duì)腔腸動(dòng)物浮浪幼蟲階段的詳細(xì)劃分具有重要意義。然而, 目前已報(bào)道的相關(guān)文獻(xiàn)均未對(duì)這一階段進(jìn)行詳細(xì)描述, 如丁耕蕪等[4]在海蜇生活史中對(duì)浮浪幼蟲的形態(tài)、顏色等進(jìn)行簡(jiǎn)單描述。此外, 在黃斑海蜇[7]、白色霞水母[8-9]、沙海蜇[10]、地中海水母(Rhopilema nomadica)[16],Rhizostoma octopus[17]的生活史中, 浮浪幼蟲也均未被詳細(xì)描述。作者分析這主要是因?yàn)槔徦父±擞紫x階段歷時(shí)短, 造成許多現(xiàn)象難易詳細(xì)觀察,如海蜇浮浪幼蟲階段最短為18 h。本研究經(jīng)過反復(fù)觀察發(fā)現(xiàn)浮浪幼蟲的縱軸是不斷伸長(zhǎng)的, 同時(shí)其活力和對(duì)外界刺激反應(yīng)也呈現(xiàn)先加速后降低的趨勢(shì), 因此作者建議將海蜇浮浪幼蟲分為早、中、晚3個(gè)階段。
3.2 四觸手螅狀體兩種形成方式中的杯狀體階段
除海月水母(Aurelia aurita)等少數(shù)可通過出芽繁殖方式形成四觸手螅狀體外, 浮浪幼蟲變態(tài)發(fā)育和足囊萌發(fā)是缽水母形成四觸手的僅有兩種方式。前期海蜇生活史的研究并未對(duì)這兩種方式進(jìn)行詳細(xì)描述。本實(shí)驗(yàn)觀察對(duì)比了這兩種螅狀體形成方式, 結(jié)果發(fā)現(xiàn)二者均形成類似于高腳酒杯狀的杯狀體階段,之后杯狀體進(jìn)一步發(fā)育, 在球形頂部四周出現(xiàn)觸手原基, 并發(fā)育成四條觸手螅狀體。Dong等[8]曾在研究白色霞水母浮浪幼蟲附著變態(tài)中指出其浮浪體囊附著后, 柄部拉長(zhǎng), 頂點(diǎn)隆起膨大呈球形, 球形托部頂端向4個(gè)對(duì)稱方向隆起, 形成早期螅狀體觸手。此外, Pitt[6]和 Littleford[18]分別在彩色水母(Catostylus mosaicus)和五卷須金黃水母(Dactylometra quinquecirrha) 的浮浪幼蟲附著變態(tài)過程中也有類似報(bào)道。因此, 作者推斷杯狀體階段是普遍存在于缽水母生活史中的。同時(shí), 由于該階段既不像浮浪幼蟲一樣能夠游動(dòng), 又不同于四觸手螅狀體, 未發(fā)育出觸手和口, 不能攝食外源性食物, 因此該階段是海蜇等缽水母類開口攝食的一個(gè)節(jié)點(diǎn), 即杯狀體階段之前是內(nèi)源自營(yíng)養(yǎng)型, 之后則為外源性營(yíng)養(yǎng)型, 作者建議將其分離出來, 作為海蜇生活史的一個(gè)單獨(dú)階段。
3.3 浮游幼體不同階段的劃分
目前對(duì)海蜇浮游幼體的劃分爭(zhēng)議主要在初生碟狀體到幼蜇這一階段。黃鳴夏等[19]根據(jù)口腕形態(tài)將碟狀體其分為四葉口腕期與八葉口腕期; 王緒峨等[20]建議按傘徑長(zhǎng)度將其分為前期碟狀體、中期碟狀體與后期碟狀體; 孫明等[11]將其分為早期碟狀體、中期碟狀體和晚期碟狀體, 同時(shí)規(guī)定晚期碟狀體是從到水母體的過渡階段, 此階段的中央口未愈合, 但已有水母體的初期形態(tài)。然而, 在生產(chǎn)和資源調(diào)查過程中, 受條件限制, 無法按上述分類方法進(jìn)行分類。經(jīng)過反復(fù)觀察, 本實(shí)驗(yàn)根據(jù)傘部中膠層變化、中央口發(fā)育狀況及棒狀附著器的出現(xiàn)與否 3個(gè)明顯的形態(tài)特征, 對(duì)海蜇浮游幼體重新進(jìn)行了劃分, 將其分為碟狀體階段、稚蜇階段和幼蜇階段, 分別描述如下: 碟狀體: 碟狀體初期從中央口到頂部是一個(gè)暢通的開放管道, 消化腔通道口尚未封閉, 在頂部中央留有一圓形洞口, 待2~3 h頂部圓形孔洞閉合后, 經(jīng)4 d左右才完全愈合; 稚蜇: 碟狀體以原愈合孔洞為中心, 傘部不斷加厚, 形成厚的中膠層, 同時(shí)4對(duì)片狀口腕順中央口下垂, 形成八葉口腕, 其中央口周圍開始分化, 出現(xiàn)小的吸口, 此時(shí)由于中央口未愈合,并隨海蜇增長(zhǎng)而增大, 其直徑可超過1 mm, 該階段的海蜇能夠攝食捕食周圍的仔稚魚, 為稚蜇前期。當(dāng)中央口完全愈合后, 稚蜇只能通過周圍吸口攝食食物, 幾乎喪失了攝食仔稚魚的能力, 該階段為稚蜇后期。幼蜇:稚蜇繼續(xù)生長(zhǎng), 傘緣逐漸下包, 由草帽狀變?yōu)榘肭驙?在愈合的口腕外端長(zhǎng)出棒狀附著器, 我們稱之為幼蜇,此時(shí)海蜇的自主游動(dòng)能力明顯增強(qiáng), 除生殖腺未發(fā)育外, 該階段海蜇與成蜇形態(tài)上無本質(zhì)差別。
綜上, 本研究在原有海蜇生活史基礎(chǔ)上, 對(duì)海蜇浮浪幼蟲到幼蟄階段的生活史進(jìn)行了補(bǔ)充研究,具體如下: 浮浪幼蟲階段進(jìn)一步細(xì)分為早期浮浪幼蟲、中期浮浪幼蟲和晚期浮浪幼蟲; 浮浪幼蟲和足囊萌發(fā)過程中加入杯狀體階段; 海蜇浮游階段劃分為碟狀體階段、稚蜇階段、幼蜇階段和成蜇階段。
[1]吳寶鈴.海蜇[J].生物學(xué)通報(bào), 1955, 4: 35-40.
[2]洪惠馨, 張士美, 王景池.海蜇的生物學(xué)[M]//海蜇,北京: 科學(xué)出版社, 1978: 1-12.
[3]Kramp P L.Synopsis of the medusae of the world[J].Jour Mar Biol Ass UK, 1961, 40: 1-469.
[4]丁耕蕪, 陳介康.海蜇的生活史[J].水產(chǎn)學(xué)報(bào), 1981,5(2): 93-104.
[5]Fautin D G.Reproduction of cnidaria[J].CanadianJournal of Zoology, 2002, 80(10): 1735-1754.
[6]Pitt K A.Life history and settlement preferences of the edible jellyfish,Catostylus mosaicus(Scyphozoa:) [J].Marine Biology, 2000, 136(2): 269-279.
[7]劉春洋, 王文波, 董婧, 等.黃斑海蜇的生活史及幾種缽水母類螅狀體形態(tài)特征的比較[J].漁業(yè)科學(xué)進(jìn)展, 2009, 30(4): 102-107.
[8]Dong J, Liu C Y, Wang Y Q, et al.Laboratory observations on the life cycle ofCyanea nozakii(Semeostomida, Scyphozoa) [J].Acta Zoologica Sinica, 2006, 52(2): 389-395.
[9]Liu C S, Zhuang Z M, Chen S Q, et al.A large-scale breeding experiment and observations on development ofCyanea nozakii(Scyphozoa, Cyaneidae) [J].Hydrobiologia, 2014, 1883-1888.
[10]Ohtsu K, Kawahara M, Ikeda H, et al.Experimental induction of gonadal maturation and spawning in the giant jellyfishNemopilema nomurai(Scyphozoa: Rhizostomeae)[J].Marine Biology, 2007, 152: 667-676.
[11]孫明, 董婧, 趙云, 等.沙蜇與海蜇晚期碟狀體的形態(tài)學(xué)研究[J].漁業(yè)科學(xué)進(jìn)展, 2012, 31(1): 48-53.
[12]曹亮, 劉金虎, 于鑫, 等.海蜇浮游幼體對(duì)褐牙鲆卵和初孵仔魚[J].海洋與湖沼, 2012, 43(3): 513-519.
[13]劉春勝, 鄒安革, 莊志猛, 等.海蜇浮浪幼蟲及早期螅狀體攝食行為的研究[J].海洋與湖沼, 2012, 43(3):545-549.
[14]宋大祥.無脊椎動(dòng)物的系統(tǒng)演化[J].生物學(xué)通報(bào),1983, 5: 9-12.
[15]Groger H, Schmid V.Larval development in Cnidaria: a connection to Bilateria [J].Genesis, 2001, 29(3):110-114.
[16]Lotan A R, Ben-Hillel, Loya Y.Life cycle ofRhopilema nomadica: a new immigrant scyphomedusan in the Mediterranean [J].Marine Biology, 1992, 112(2): 237-242.
[17]Holst S, S?tje I, Tiemann H, et al.Life cycle of the rhizostome jellyfishRhizostoma octopus(L.) (Scyphozoa, Rhizostomeae), with studies on cnidocysts and statoliths[J].Marine Biology, 2007, 151(5): 1695-1710.
[18]Littleford R A.The life cycle ofDactylometra quinquecirrha, L.Agassiz in the Chesapeake Bay[J].Biol Bull,1939, 77(3): 368-381.
[19]黃鳴夏, 王永順, 孫忠.溫度和鹽度對(duì)海蜇碟狀體生長(zhǎng)及發(fā)育的影響[J].浙江水產(chǎn)學(xué)院學(xué)報(bào), 1987, 6(2): 105-110.
[20]王緒峨, 宋向軍, 馬建新, 等.海蜇幼體與對(duì)蝦幼體相食關(guān)系的實(shí)驗(yàn)觀察[J].動(dòng)物學(xué)雜志, 1995, 30(6): 30-33.
