壁虎在垂直的不同材料表面的運動與附著行為研究
2014-12-05 06:53:56吉愛紅汪中原王周義戴振東
中國機械工程 2014年4期
吉愛紅 顧 偉 汪中原 倪 勇 王周義 戴振東
南京航空航天大學,南京,210016
0 引言
在億萬年的生存與演變過程中,生物的形態結構、運動機理和行為方式逐漸與其生存環境相適應[1]。壁虎[2-3]、昆蟲[4]等能夠在各種崎嶇不平的表面或光滑的表面上自由爬行或附著。動物在不同材質表面的附著機制及運動行為的研究,對研制具有全空間自由運動能力的仿生機器人有重要意義[5-6]。
壁虎腳趾上長有數以百萬計的剛毛,在光滑壁面上,主要依靠腳趾剛毛與運動表面產生的黏附力附著于各種表面,其腳掌的黏附過程就是剛毛與運動表面黏附及脫附的過程。研究人員希望通過揭示壁虎的運動規律和附著機制,為設計仿生爬壁機器人提供依據。Dellit[7]發現壁虎在不同的大氣壓力環境下以及在充滿正負相異電荷的空氣中都依然能夠保持出色的附著能力,因此排除了壁虎依靠真空吸附或靜電引力實現附著的假設。Simmermacher[8]對壁虎腳掌進行解剖,沒有發現能夠分泌黏液的腺體,說明壁虎剛毛不像螞蟻那樣基于濕黏附機制形成附著能力。Autumn等[9-12]從壁虎的運動行為、運動力學以及壁虎腳掌的微觀結構、附著機理、附著力等方面對壁虎進行了細致的研究,測量了單根剛毛的黏附力,提出了壁虎附著基于范德華力作用的結論。Huber等[13]通過試驗研究了在納米尺度上毛細力對壁虎剛毛的附著能力的貢獻。Puthoff等[14]從材料性能改變的角度解釋了濕度對壁虎剛毛黏附力的影響。而Zaaf等[15]發現一種適應于地面生活的壁虎(eublephairs macularius)優先通過改變步距改變速度。
綜上所述,國內外學者對壁虎運動力學、附著機制開展了大量的研究,但對壁虎在不同材料表面上的運動以及壁虎腳掌附著能力與材料之間的相關性研究尚未見報道。Hiller[16-18]和 Autumn等[19]發現壁虎的黏附性能在某種程度上由表面能決定,但并不清楚是何種關系。本文測量了壁虎在不同材料的垂直表面上運動時的三維接觸反力,并同步記錄了其運動步態信息,分析討論了其附著機制。研究結果為仿壁虎機器人的設計與控制提供了仿生依據。
1 試驗對象與方法
1.1 試驗對象
試驗動物為大壁虎(gekko gecko)[20],俗名蛤蚧,購于廣西壯族自治區貴港市真真蛤蚧養殖場。為保持大壁虎的活性,實驗室專門建有可模擬大壁虎在廣西野外生存環境的壁虎養殖室,具備自然光照條件,環境溫度常年為27±2℃,濕度為70%~80%。大壁虎用水、葡萄糖和蝗蟲等喂食。試驗用大壁虎共7只,每只質量為62.9±8.1g,體長為136.0±10.0mm。在養殖室內放置可模擬試驗裝置的通道,以訓練壁虎使其適應試驗裝置通道爬行。
1.2 試驗方法
1.2.1動物運動反力與行為測試
大壁虎試驗系統如圖1所示。壁虎在垂直的不同材料表面的三維運動反力與行為測試用本實驗室自主研制的動物全空間運動反力-行為測試系統(專利號:ZL200810156169.0)[21-22]。系統包括測力平臺、信號調理與數據采集設備(SCXI系列,NI公司,美國)、高速攝像機(Mikrotron,MC1311,德國)和一套在Labview(NI公司,美國)環境下開發的可進行力學與圖像數據采集、分析與測試的軟件(圖1a)。四只mN級三維力傳感器[23](圖1b)支撐承力板形成測力平臺。根據試驗要求,承力板材料可更換為玻璃、有機玻璃、鋁、不銹鋼等。承力板嵌入兩端開口的透明的亞克力材料通道內,并與通道底面共面,形成壁虎爬行試驗通道,使壁虎在通道內保持近似直線運動。試驗通道兩側安裝有與承力表面成45°夾角的鏡子,高速攝像機(215幀/秒)從試驗通道的正上方可以同時記錄通道內及兩側鏡子內壁虎的運動行為,從而得到壁虎的三維運動行為。壁虎在垂直通道中向上爬行時,前足先踩上承力表面,接著整個身體通過承力表面,直至后足完全離開。由動物全空間運動反力-行為測試系統可得到前足的單足接觸反力、整體接觸反力和后足的單足接觸反力。4只三維力傳感器共12個通道的力信號采集與高速攝像機的圖像記錄通過外部觸發信號實現同步。
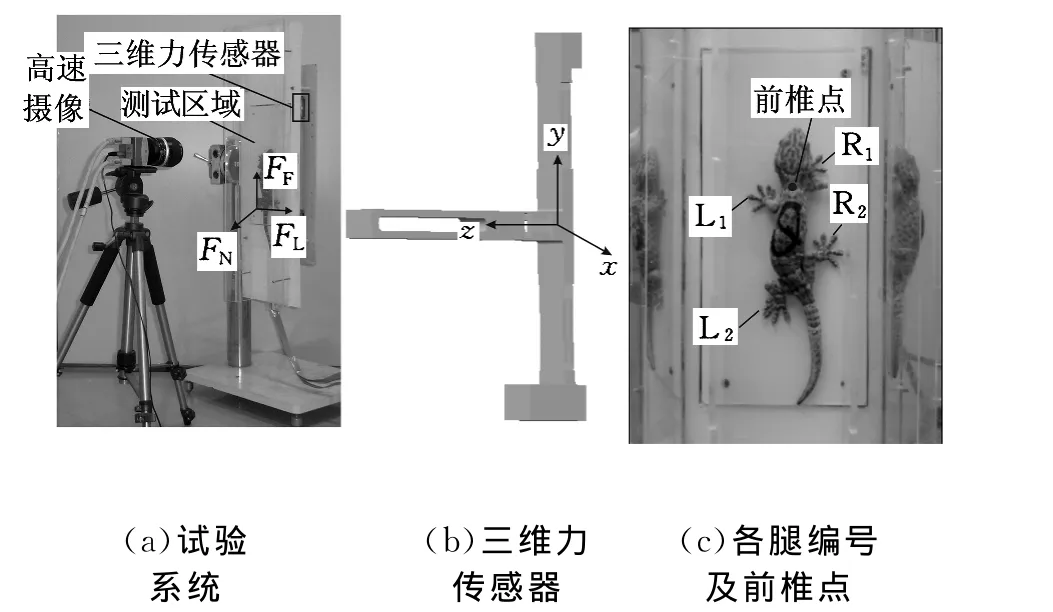
圖1 大壁虎試驗系統圖
1.2.2材料表面粗糙度與接觸角測量
用表面粗糙度輪廓儀(JB-5C,上海泰明光學儀器有限公司)測出玻璃、有機玻璃、鋁和不銹鋼等材料表面的粗糙度。用光學接觸角儀(CAM200系列,KSV儀器公司,芬蘭)測出各個表面和壁虎剛毛的接觸角,用兩液法計算得到玻璃、有機玻璃、鋁、不銹鋼和壁虎腳趾剛毛的表面能(表1)。
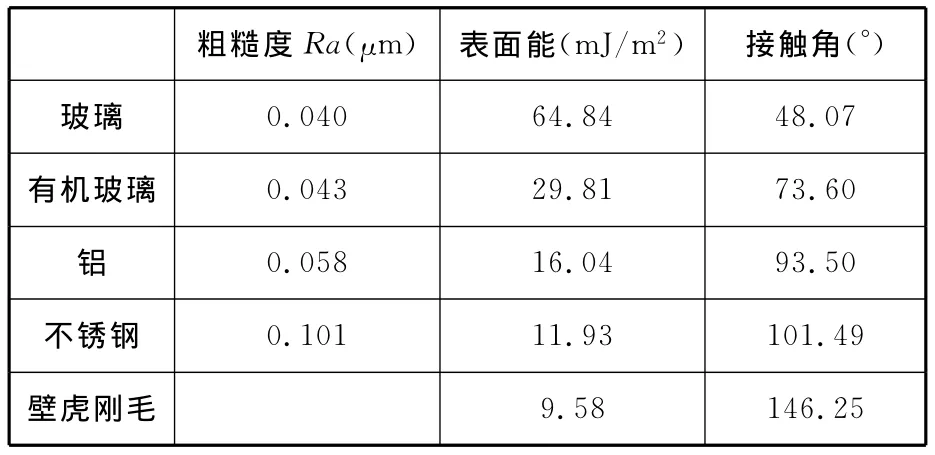
表1 不同材料的表面參數
1.2.3數據提取與分析
在記錄的高速圖像中,用定點坐標獲取軟件(Sigma Scan Pro,SPSS公司,美國)確定壁虎頸部前椎點(圖1c)的坐標,由前椎點在多幅連續圖像中移動的距離除以相應幀數圖像對應的時間可以計算出壁虎在爬行過程中的速度。為便于試驗結果的統計與分析,將壁虎的四足分別定義為L1(左前足)、L2(左后足)、R1(右前足)和 R2(右后足)(圖1c)。根據高速攝像獲取的圖片信息,判定壁虎身體與承力表面的相對位置,讀取壁虎腳掌與承力表面之間的接觸情況,并截取各傳感器所測得的力數據,從而獲得壁虎運動反力在3個方向上的分量(法向力FN,軸向力FF,側向力FL)。
試驗過程中,壁虎的體重、身體活性狀態以及環境因素等,都會對壁虎的運動反力與運動行為產生影響。因此,由大量試驗得到的壁虎運動力學與運動行為數據的統計和分析必須通過統計學工具進行。我們對不同試驗組試驗結果的差異性采用t檢驗的方法進行檢驗。當概率P>0.05時,反映了這兩組試驗數據之間沒有顯著性差異,反之則存在著差異;對于兩組或多組試驗數據的線性回歸分析則采用方差分析(ANOVA)檢驗其相關性,當概率P<0.05時,說明線性相關性顯著,反之則認為沒有顯著相關性。本文試驗數據中給出的統計值為平均值±標準偏差。
2 試驗結果
2.1 運動步態
壁虎在垂直的玻璃、有機玻璃、鋁表面運動時,L1與R2,R1與L2的步態基本一致,壁虎的身體具有左右對稱性,將左右腿的試驗參數結果(步距、步頻和占空比等)作統計檢驗,結果表明左右兩側對應腿的各參數無顯著差異性。因此,本文對試驗數據分析時將左前腿與右前腿、左后腿與右后腿的試驗數據分別進行合并分析,得到了壁虎在不同材料的垂直表面上的速度、步頻、步距、占空比等步態參數(表2)。
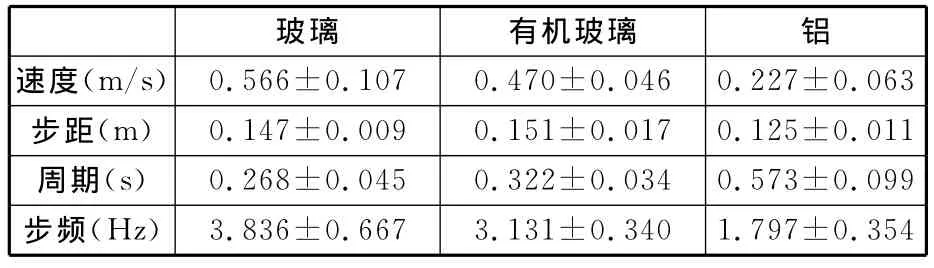
表2 壁虎在垂直的不同材料表面的運動步態參數
壁虎一個步態周期可分為兩個相位,即支撐相和擺動相。支撐相指壁虎各腳掌接觸壁面的時間,擺動相指壁虎腳掌離開壁面向前(上)邁步到再次落地之間的時間。根據壁虎腳掌與壁面接觸狀態,支撐相又分為三個階段:首先是腳掌掌心與承力板接觸,將腳趾伸直后內收使其貼附到承力板上以產生黏附力,即黏附階段;接著腳掌進入與承力板完全接觸的狀態,即附著階段;此后壁虎腳趾向外展開離開承力板,即脫附階段。壁虎在一個周期中支撐相(黏附、附著、脫附)與擺動相占整個周期的百分比見表3。
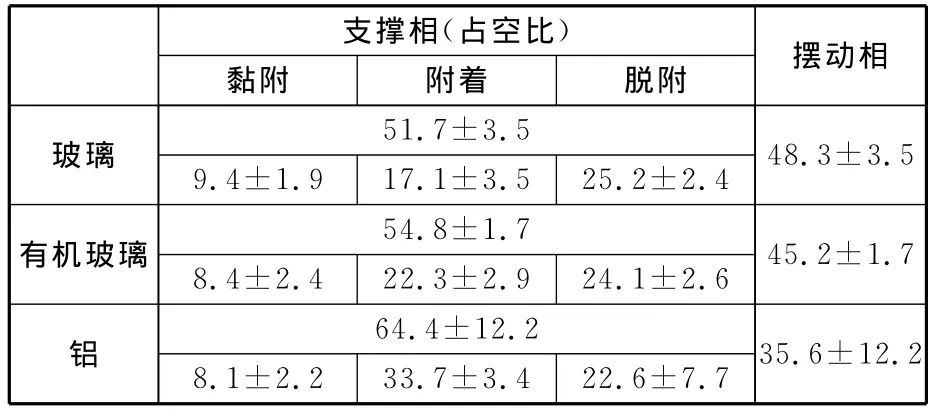
表3 壁虎在垂直的不同材料表面的一個運動周期內占空比分析 %
2.1.1玻璃表面
壁虎在垂直的玻璃表面向上爬行(圖2)時,速度為(0.566±0.107)m/s,步頻為(3.836±0.667)Hz,步頻與速度有顯著相關性(ANVOA,線性度R2=0.875,P=0.000<0.05,判定臨界值F=76.86,數據自由度df=12)。步距為(0.147±0.009)m,步距與速度亦有顯著相關性(ANVOA,R2=0.480,P=0.009<0.05,F=10.14,df=12),腳掌占空比為0.517±0.035,占空比與速度同樣也有顯著相關性(ANVOA,R2=0.082,P=0.002<0.05,F=15.29,df=12)。
在一個周期中,腳掌黏附時間為(25.4±0.49)ms,占整個周期的(9.4±1.9)%;腳掌附著時間為(45.9±1.62)ms,占整個周期的(17.1±3.5)%;腳掌脫附時間為(67.8±1.66)ms,占整個周期的(25.2±2.4)%;腳掌在空中的擺動時間為(130.2±4.49)ms,占 整 個 周 期 的 (48.3±3.5)%。
2.1.2有機玻璃表面
壁虎在垂直的有機玻璃表面向上爬行時速度為(0.470±0.046)m/s,步頻為(3.131±0.340)Hz,步頻與速度有顯著相關性(ANVOA,R2=0.360,P=0.016<0.05,F=3.379,df=7),步距為(0.151±0.017)m,步距與速度無顯著的相關性(ANVOA,R2=0.302,P=0.150,df>0.05,F=2.595,df=7),腳掌占空比為0.548±0.017,占空比與速度有顯著相關性(ANVOA,R2=0.066,P=0.049<0.05,F=0.425,df=7)。
在一個周期中,腳掌黏附時間為(17.6±0.44)ms,占整個周期的(8.4±2.4)%;腳掌附著時間為(45.2±1.31)ms,占整個周期的(22.3±2.9)%;腳掌脫附時間為(48.9±1.25)ms,占整個周期的(24.1±2.6)%;腳掌在空中的擺動時間為 (91.7±1.57)ms,占 整 個 周 期 的 (45.2±1.7)%。
2.1.3鋁表面
壁虎在垂直的鋁表面向上爬行時速度為(0.227±0.063)m/s,步頻為(1.797±0.354)Hz,步頻與速度有顯著相關性(ANVOA,R2=0.926,P=0.000<0.05,F=74.65,df=8),步距為(0.125±0.011)m,步距與速度亦有顯著相關性(ANVOA,R2=0.640,P=0.017<0.05,F=10.672,df=8),腳掌占空比為0.641±0.012,占空比與速度同樣也有顯著相關性(ANVOA,R2=0.293,P=0.046<0.05,F=2.484,df=8)。
在一個周期中,腳掌黏附時間為(46.3±1.01)ms,占整個周期的(8.1±2.2)%;腳掌附著時間為(193.1±6.54)ms,占整個周期的(33.7±3.4)%;腳掌脫附時間為(129.3±9.91)ms,占整個周期的(22.6±7.7)%;腳掌在空中的擺動時間為(205.8±25.2)ms,占 整個 周 期 的 (35.6±12.2)%。
2.1.4不銹鋼表面
將壁虎放在垂直的不銹鋼壁面上,壁虎會直接掉落到地面。因此壁虎不能附著在垂直的不銹鋼表面,更不能在不銹鋼壁面上向上自如爬行。
2.2 接觸反力
2.2.1整體接觸反力
在垂直的玻璃、有機玻璃和鋁這三種材料表面上,壁虎向上爬行時不但要抵消身體的重力,還要推動身體向上前進,軸向力FF始終保持為前進方向。因為一個周期內壁虎的運動是一個加減速運動過程[12],所以軸向力FF大小在體重上下波動。壁虎的運動也是支撐相和擺動相交替轉換的過程,與承力板垂直的法向力FN以及與運動方向垂直的側向力FL也隨之發生方向上的交替變化(圖2),即正負值波動變化。由試驗結果(表4)可知,壁虎在玻璃、有機玻璃和鋁表面上向上爬行時的三維運動反力分量即軸向力FF、法向力FN和側向力FL的最大值都依次減小。從而可以得到壁虎在垂直的不同材料表面上運動時,在玻璃表面上的運動反力最大,在鋁表面上的運動反力最小。
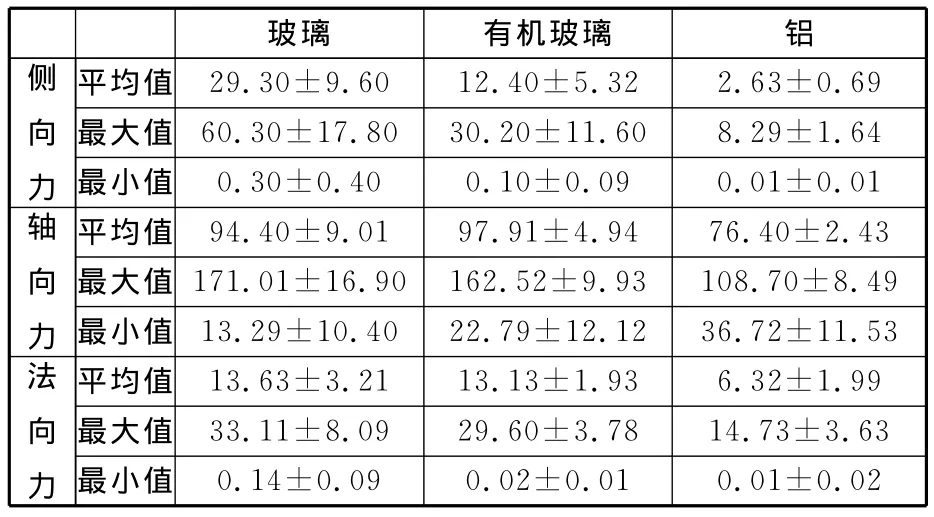
表4 壁虎在垂直的不同材料表面整體接觸反力占體重的百分比 %
2.2.2單足接觸反力
如前所述,采用對角運動步態的壁虎在垂直的通道內爬行時,在進入承力板的初始階段,一只前腳掌與承力板接觸;在身體離開承力板的最后階段,一只后腳掌與承力板接觸。由動物全空間運動反力-行為測試系統可以測試單只腳掌與承力板接觸時的三維接觸反力(圖3)。由試驗結果可知,與整體接觸反力不同,在玻璃、有機玻璃和鋁表面上,壁虎的單足軸向力FF、法向力FN和側向力FL的大小和方向都沒有明顯差異。
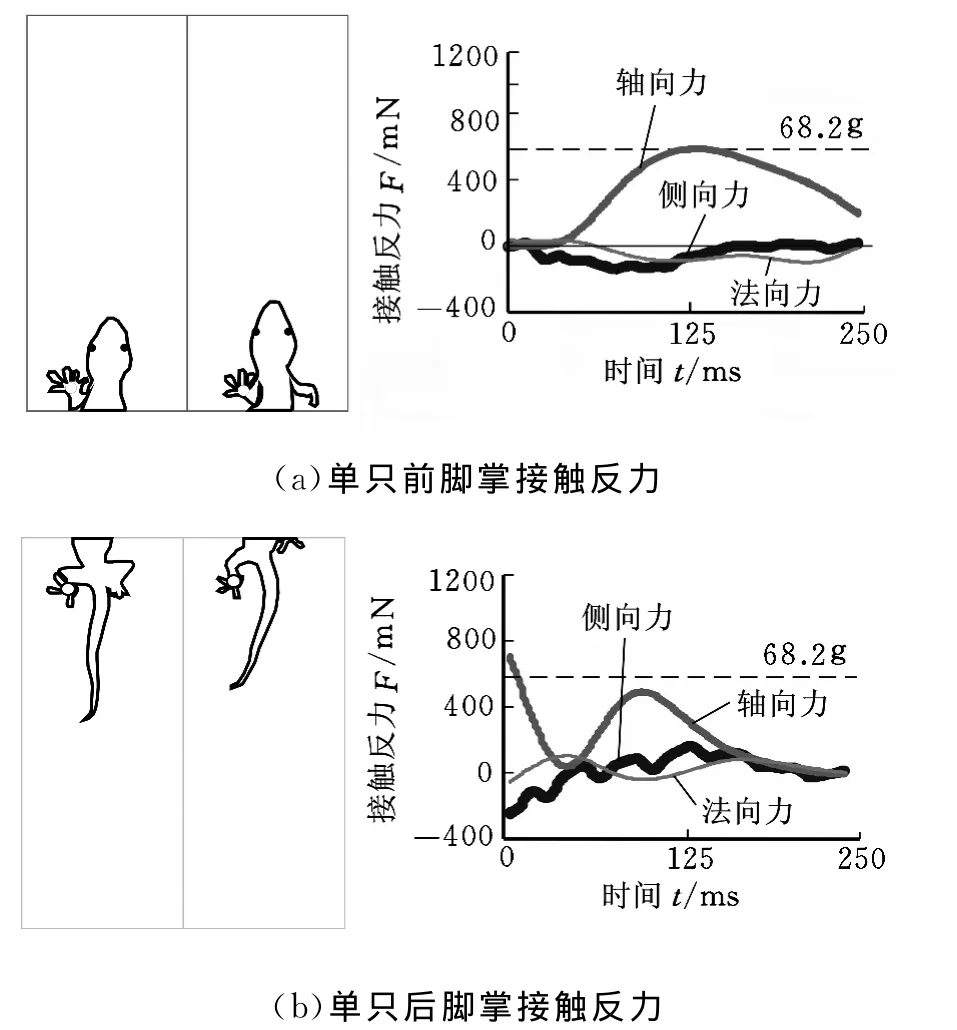
圖3 大壁虎單只腳掌與承力板間的接觸反力
前腿和后腿的軸向力方向都始終為壁虎向上運動的方向。在一個對角步態周期中,軸向力大部分時間都小于自身的體重。但一條前腿和一條后腿(如左前腿和右后腿)的軸向力合力大小在體重值附近呈周期性上下波動,這與測得的整體接觸反力的軸向力分量結果一致。前腿對承力板的法向力指向壁虎身體,即前腿產生的黏附力使身體附著在垂直表面;后腿對承力板的法向作用力的方向則與前腿相反,與承力板之間產生了推力。壁虎單腿對承力板的側向力的方向始終垂直于運動軸向并指向身體。側向力的最大值出現在支撐相的附著階段。在側向力的作用下,壁虎的身體盡可能地橫向伸展并貼附垂直的運動表面。
3 討論
3.1 運動步態
試驗結果表明,壁虎在垂直的不同材料表面向上爬行時,采用對角步態以較快的速度運動。以一側的前腿與另一側的后腿形成一組,兩組的運動相互交替實現擺動相與支撐相的連續快速更替。
在玻璃、有機玻璃和鋁表面上,壁虎均能保持穩定的連續爬行;在不銹鋼表面不能附著,也不能向上爬行,直接從垂直的不銹鋼表面下滑甚至掉落地面。在玻璃、有機玻璃和鋁表面上的平均速度依次減小。在玻璃上爬行的平均速度是鋁表面爬行平均速度的兩倍。
在玻璃和鋁表面,速度與步頻和步距都有顯著相關性,說明在玻璃和鋁表面,爬行速度隨著步頻和步距的同時增大而提高,即壁虎通過同時增大步距和提高步頻來相應提高向上爬行的速度。在有機玻璃表面,速度與步頻有顯著相關性,與步距無相關性,說明速度隨著步頻增加而提高,即壁虎通過加快運動頻率提高向上爬行的速度。
壁虎在垂直的三種材料表面向上爬行時的一個運動周期內占空比均大于0.5,充分保證了運動時的可靠附著與穩定性。在玻璃、有機玻璃和鋁表面上,壁虎一個運動周期的占空比依次增大,即在相應表面上處于支撐相的時間依次增加。在三種表面上,速度與占空比都存在顯著相關性,且速度和占空比隨著周期內在空中的擺動時間的增加而提高。在一個運動周期內處于支撐相的三個階段,鋁表面在附著階段花費的時間最長,玻璃表面花費的時間最短。從運動與附著的安全性角度來分析,壁虎在玻璃表面上運動時最安全,與鋁表面相比,不需要花費較多的時間用于附著。可以認為,在玻璃、有機玻璃、鋁以及不能運動與附著的不銹鋼表面,壁虎運動的安全與穩定性依次降低。
因此,壁虎在垂直的不同材料表面上運動時,隨著運動表面材料的不同,壁虎的運動步態會發生明顯的變化。在相對安全的表面,壁虎運動較快,通過增大步距或提高步頻來提高速度。在附著性能較弱的表面,壁虎會降低爬行速度以保證運動的穩定與安全。步距明顯減小的同時,單位時間內的邁步頻率也顯著降低。在一個運動周期內,壁虎還會增大占空比,在支撐相的三個階段顯著增大附著時間,縮短黏附、脫附時間以及腿在空中的擺動時間,提高在附著性能較弱表面上運動與附著的安全性和穩定性。
3.2 運動力學
如前所述,試驗測得壁虎在玻璃、有機玻璃和鋁表面上向上爬行時的三維運動反力分量即軸向力FF、法向力FN和側向力FL的最大值都依次減小,因此壁虎在玻璃、有機玻璃和鋁表面上向上爬行時的運動反力也依次減小,壁虎在三種材料上的運動速度也依次減小,由此可知壁虎在附著安全性較高的表面運動時,消耗功率較大。但是由于壁虎在三種表面上的運動周期依次增大,所以在一個運動周期內,經計算可知,壁虎在不同的材料表面運動時做功基本相等。在一個運動周期內,壁虎的運動是減速和加速的交替變換過程,而在整個運動過程中,壁虎的運動則是近似勻速向上的直線運動。因此壁虎的做功主要用于壁虎向上運動時勢能的增加(圖2e)。
3.2.1軸向力
壁虎向上爬行時的軸向力作為運動的驅動力,始終與重力方向相反。軸向力的大小在重力上下波動。軸向力與重力的合力為正時,壁虎加速向上爬行;經過半個周期后,軸向力與重力的合力變為負值,壁虎減速運動。
3.2.2法向力
整體法向接觸反力在壁虎運動過程中發生方向上的交替變化。壁虎的腳掌離開壁面時,腳掌上的剛毛與壁面間產生脫附行為,法向力為正;壁虎腳掌接觸到壁面,腳掌剛毛與壁面間產生黏附行為,法向力為負值。向上爬行時近似勻速運動的壁虎所受的摩擦力大致與體重相當,由摩擦力、法向力和剛毛與壁面間的摩擦因數關系可知,正負法向力的最大值基本相等(圖2c)。壁虎通過前腿產生的黏附力使身體附著在垂直表面,而后腿法向力方向與前腿的相反。以前腿與壁面接觸點為支點,由后腿法向力產生的力矩可以平衡壁虎身體重心偏離壁面所產生的傾覆力矩。
3.2.3側向力
壁虎作為四足匍匐動物,在向上爬行過程中,在方向交替變化的側向力的作用下身體發生柔性彎曲,質心橫向左右擺動,在對角步態下的身體質心始終保持在前后對側兩條支撐腿連成的對角線附近,從而保持運動的穩定性。這與身體軀干呈剛性的六足昆蟲或八足蜘蛛不同,六足或八足動物多采用三角步態[24],其質心始終位于由三條支撐腿或更多腿組成的三角形內以保持運動的穩定性。壁虎在垂直壁面上運動時各條腿受到的側向力使身體橫向伸展而盡可能地貼附在垂直壁面上。作為匍匐動物,壁虎在垂直壁面上運動時的各條腿上的側向力可以保持運動時的穩定性。
3.3 附著機制
由壁虎在垂直的不同材料表面運動行為與運動反力的試驗結果分析可知,在玻璃、有機玻璃、鋁和不銹鋼表面,壁虎的運動與附著能力依次降低。壁虎腳掌剛毛約2~10μm 寬,100μm 長[13],剛毛的尺寸遠遠大于各材料的表面粗糙度(表1),因此不能從粗糙度的角度來分析壁虎在不同材料表面上的附著性能。而且壁虎也不可能通過腳爪的機械鎖合黏附來實現附著。壁虎剛毛的接觸角為146.25°,表現為強疏水性,因此也不可能通過毛細力作用實現黏附。進一步驗證了Autumn提出的壁虎腳掌剛毛基于范德華力作用實現黏附的結論。由表1可知,玻璃、有機玻璃、鋁和不銹鋼的表面能依次降低,由此可知,壁虎在垂直表面的運動與附著能力與材料的表面能有關,隨著接觸面材料表面能的增大而提高。
4 結語
大壁虎在垂直表面上爬行以對角步態為主,主要通過提高步頻來提高速度。這與通過改變步距而改變速度的適應于地面生活的壁虎(eublephairs macularius)不同。這是不同屬的壁虎分別對地面和壁面生活長期進化適應的結果。
壁虎在垂直的玻璃、有機玻璃、鋁和不銹鋼表面的運動與附著能力依次降低。壁虎腳掌剛毛附著基于范德華力作用機制。壁虎在垂直表面的運動與附著能力隨著接觸面材料的表面能的增大而提高。
上述研究可對基于仿剛毛陣列黏附的仿生爬壁機器人腳掌黏附能力提升及其與接觸材料表面適應性的設計,提供參考依據。
[1]畢樹生,宗光華.關于21世紀初我國仿生機械與仿生制造的若干思考[J].中國機械工程,2001,12(10):1201-1204.Bi Shusheng,Zong Guanghua.Some Ideas about Bionic Machines and Bionic Manufacture of Our Country in the Early 21st Century[J].China Mechanical Engineering,2001,12(10):1201-1204.
[2]戴振東,于敏,吉愛紅,等.動物驅動足摩擦學特性研究及仿生設計[J].中國機械工程,2005,16(16):1201-1204.Dai Zhendong,Yu Min,Ji Aihong,et al.Study on Tribological Characteristics of Animals’Driving Pads and Their Bionic Design[J].China Mechanical Engineering,2005,16(16):1201-1204.
[3]王周義,王金童,吉愛紅,等.大壁虎在天花板表面的運動行為與動力學研究[J].科學通報,2010,55(9):841-848.Wang Zhouyi,Wang Jintong,Ji Aihong,et al.Gecko’s Movement Behavior and Dynamics Research on Ceiling Surface[J].Chinese Sci.Bull.,2010,55(9):841-848.
[4]Frantsevich L,Ji A H,Dai Z D,et al.Adhesive Properties of the Arolium of a Lantern-fly,Lycorma Delicatula[J].J.Insect Physiol.,2008,54:818-827.
[5]吉愛紅,戴振東,周來水.仿生機器人的研究進展[J].機器人,2005,27:284-288.Ji Aihong,Dai Zhendong,Zhou Laishui.Research Development of Bio-inspired Robotics[J].Robot,2005,27:284-288.
[6]張昊,龍瑋潔,李錦方,等.一種仿壁虎機器人側向地壁過渡方式及步態[J].中國機械工程,2013,24(5):573-579.Zhang Hao,Long Weijie,Li Jinfang,et al.Transition Gait from Ground to Wall for a Gecko-mimicking Robot[J].China Mechanical Engineering,2013,24(5):573-579.
[7]Dellit W F.Zur Anatomie and Physiologie der Geekozehe[J].Jena.Z.Naturw,1934,68:613-656.
[8]Simmermacher G.Untersuchungen uber Haftapparate an Tarsalgliedern von Insekten [J].Zeitschrift far Wissenschaftliche Zoologie,1884,40:481-556.
[9]Autumn K,Liang Y A,Hsieh S T,et al.Adhesive Force of a Single Gecko Foot-hair[J].Nature,2000,405:681-684.
[10]Autumn K,Sitti M.Evidence for van der Waals Adhesion in Gecko Setae[J].PANS,2002,99:11993-12502.
[11]Autumn K,Hsieh S T,Dudek D M,et al.Dynamics of Geckos Running Vertically[J].J.Exp.Biol.,2006,209:260-272.
[12]Chen J J,Peattie A M,Autumn K,et al.Differential Leg Function in a Sprawled-posture Quadrupedal Trotter[J].J.Exp.Biol.,2006,209:249-259.
[13]Huber G,Mantz H,Spolenak R,et al.Evidence for Capillarity Contributions to Gecko Adhesion from Single Spatula Nanomechanical Measurements[J].PANS,2005,102:16293-16296.
[14]Puthoff J B,Prowse M S,Wilkinson M,et al.Changes in Materials Properties Explain the Effects of Humidity on Gecko Adhesion[J].J.Exp.Biol.,2010,213:3699-3704.
[15]Zaaf A,van Damme R,Herrel A,et al.Spatiotemporal Gait Characteristics of Level and Vertical Locomotion in a Ground-dwelling and a Climbing Gecko[J].J.Exp.Biol.,2001,204:1233-1246.
[16]Hiller U.Untersuchungen zum Feinbau und zur Funktion der Haftborsten von Reptilien[J].Z.Morphol.Tiere,1968,62:307-362.
[17]Hiller U.Correlation between Corona-discharge of Polyethylene-films and the Adhering Power of Tarentola M.mauritanica(Rept.)[J].Forma et Functio,1969,1:350-352.
[18]Hiller U.Comparative Studies on the Functional Morphology of Two Gekkonid Lizards[J].J.Bombay Nat.Hist.Soc.,1975,73:278-282.
[19]Autumn K,Peattie A.Mechanisms of Adhesion in Geckos[J].Int.Comp.Biol.,2002,42,1081-1090.
[20]中國野生動物保護協會.中國爬行動物圖鑒[M].鄭州:河南科學技術出版社,2002.
[21]Dai Z D,Wang Z Y,Ji A H.Dynamics of Gecko Locomotion:a Force-measuring Array to Measure 3DReaction Forces[J].J.Exp.Biol.,2011,214:703-708.
[22]吉愛紅,戴振東,顏化冰,等.動物運動力學測試系統[J].傳感器與微系統,2006,25(12):59-61.Ji Aihong,Dai Zhendong,Yan Huabing,et al.Study on Animal’s Locomotive Mechanics Measurement System[J].Transducer and Microsystem Technologies,2006,25(12):59-61.
[23]吳強,俞志偉,吉愛紅,等.一種小型電阻應變式3維力傳感器的仿真設計[J].中國機械工程,2011,22(11):1288-1293.Wu Qiang,Yu Zhiwei,Ji Aihong,et al.Development of a Miniature Resistance Strain Three Dimensional Force Sensor[J].China Mechanical Engineering,2011,22(11):1288-1293.
[24]Wang Z Y,Wang J T,Ji A H,et al.Movement Behavior of a Spider on a Horizontal Surface[J].Chinese Sci.Bull.,2011,56:2748-2757.
