分蘗期水分脅迫對不同栽培方式水稻生長發育及產量的影響
2014-12-05 02:57:40鄭華斌劉建霞
作物研究 2014年5期
姚 林,鄭華斌,劉建霞,賀 慧,黃 璜*
(1湖南農業大學生物科學與技術學院,長沙410128;2農業部華中地區作物栽培科學觀測實驗站,湖南長沙410128;3湖南省作物多熟制工程技術研究中心,長沙410128;4湖南農業大學農學院,長沙410128)
水稻在不同的生育階段對水分的需求不一,承受一定程度的水分脅迫后其反應也不相同,不同生育期對其敏感性不同,在反應敏感的階段進行水分脅迫,會造成光合作用減弱,生長發育受阻,甚至嚴重影響產量[1~5]。而水稻全生育期中,分蘗期對水分反應遲鈍、抗旱性強,節水潛力大[6]。
壟作梯式栽培是將傳統的淹水栽培改為半濕潤栽培的一種種植方式,改平作為壟作,改變了田間原有地形,加厚了水稻生長的熟土層,地下水位相對降低,擴大了水稻的營養面積,有利于水稻根系的生長[7];增加了群體有效受光面積,能充分利用邊際效應,優化群體結構,協調群體與個體間的矛盾,增強田間通風透光性,降低土壤濕度,提高土壤溫度,有利于有益微生物的繁殖,有效養分增加,從而增加分蘗數、成穗率、結實率和千粒重[8]。壟作增加入滲,提供貯水源泉,壟作下土壤孔隙發育較好而入滲較大。此外壟作栽培可有效防止水土流失,增加貯水載體。壟作栽培提高作物對土壤水分的利用率,壟體表面土層疏松,而壟體下層緊實,上虛下實的耕層有利于調節土壤水分。水稻在改平作為壟作后,徹底改變了淹水平作時以重力下滲為主的水分運動形式,溝內始終保持著穩定而又可調的水層,在土壤毛細管引力和吸水力的作用下,壟溝內的水分源源不斷地輸向壟頂,使壟體土壤較穩定地保持毛管水狀況,有利于水分的輸送[9,10]。
前人對壟作栽培和水分脅迫對水稻影響單獨研究的較多,但有關不同栽培方式下水稻進行水分脅迫處理對植株耗水量、水分利用效率的影響研究較少。本試驗研究了不同栽培方式下,分蘗期水分脅迫對水稻的生長特性、產量和水分利用效率的影響,旨在探討在壟作梯式栽培下,水稻對土壤水分的適應性,以期為水稻的節能節水高產栽培提供技術依據。
1 材料與方法
1.1 試驗材料
試驗于2013年6月開始,在湖南省農科院試驗大棚內進行盆栽試驗。供試土壤為第四紀紅色粘土發育的紅黃泥水稻土,土壤有機質33.51 g/kg,全氮1.52 g/kg,全磷0.94 g/kg,全鉀12.68 g/kg,水解氮120.12 mg/kg,有效磷34.78 mg/kg,速效鉀130.71 mg/kg。供試水稻材料為湘晚秈13號。
1.2 試驗方法
隨機區組設計。試驗設2種栽培方式處理和4種水分處理,各3次重復,每個重復7盆。栽培方式為傳統栽培(對照,C1)和壟作栽培(C2);水分脅迫處理分別為對照(W1:灌水深度5 cm,C2的灌水量與C1相同)、輕度脅迫(W2:灌溉量為 W1的70%)、中度脅迫(W3:灌溉量為W1的50%)、重度脅迫(W4:灌溉量為W1的30%)。當C1W1中的水層低于1 cm時,再灌水至5 cm,其它處理同時定量灌溉。
采用盆栽試驗方法。盆的規格為直徑28 cm,高33 cm,盆底有直徑為1 cm的小圓孔。每盆裝風干土10 kg,在盆內模擬起壟,壟高15 cm,寬20 cm。施肥按 N 180 kg/hm2、P2O575 kg/hm2、K2O 150 kg/hm2計算,其中P以底肥一次性施入,K按基、蘗肥各50%分2次施入,N按基肥∶蘗肥∶穗肥=5∶3∶2分3次施入。6月17日播種,7月9日移栽,每盆栽2叢,株距22 cm。移栽后12 d開始進行水分脅迫處理,共脅迫25 d。8月15日脅迫結束時,取樣測定葉面積與根系特性。
1.3 測定項目與方法
(1)分蘗動態。移栽后12 d開始,定點6叢觀察水稻分蘗情況,每隔3 d觀測一次植株的分蘗數。
(2)葉面積指標。取4株代表性植株,直接測量葉長與寬,葉面積=長×寬×0.75。
(3)根系特性測定。取樣2株,測定單株總根數、最長根長、白根數、根體積(排水法)和根直徑、根系干重和地上部干重;根系吸收面積和活躍吸收面積的測定:用甲烯藍法[7];根系活力測定用α-萘胺法[8]。
(4)產量。谷粒成熟時,取5盆測定水稻經濟性狀,包括單株有效穗數、每穗總粒數、每穗實粒數、每穗空粒數和千粒重。
2 結果與分析
2.1 水分脅迫對水稻分蘗的影響
從圖1可知,兩種栽培方式下,不同水分處理對水稻分蘗數量均有影響,且影響程度與水分脅迫一致,即輕度水分脅迫對水稻分蘗影響不大,中度、重度水分脅迫則嚴重降低分蘗數。壟作梯式栽培下水稻的分蘗數及分蘗率均高于傳統栽培,但差異不顯著。
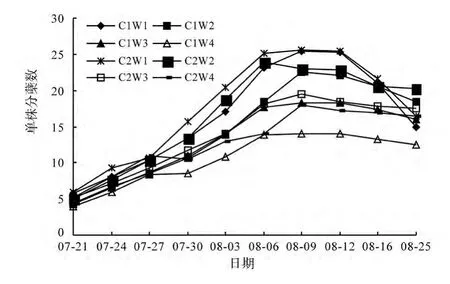
圖1 各處理的水稻分蘗動態Fig.1 Tillering dynamics of rice in different treatments
2.2 水分脅迫對水稻成穗率和葉面積的影響
栽培方式和水分脅迫對水稻成穗率有顯著影響,表現為C2>C1,W4>W3>W2>W1,同時 C因素(栽培方式)和W因素(水分處理)之間的互作效應對水稻成穗率影響顯著(表1)。
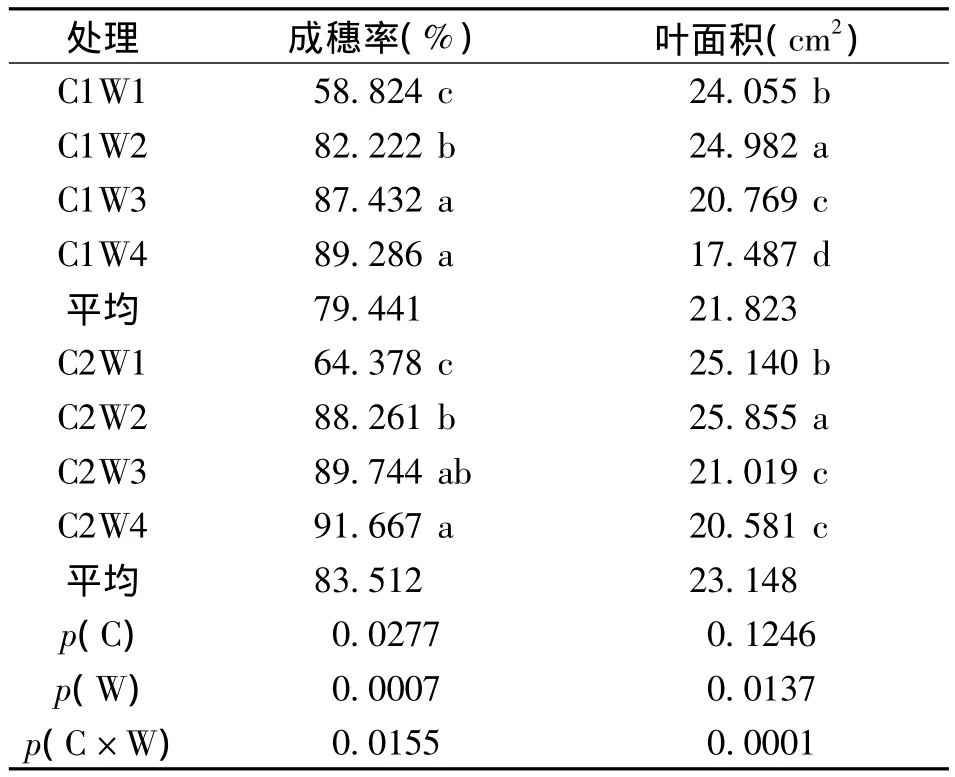
表1 分蘗期水分脅迫處理的水稻成穗率及葉面積差異Table 1 Difference in earing rate and leaf area of rice among water stress treatments
水分脅迫會制約水稻葉面積的擴展,隨著水分脅迫的加重,葉面積降低。從表1可見,水分脅迫對水稻葉面積影響顯著,表現為W2>W1>W3>W4,栽培方式對葉面積的影響不顯著,但栽培方式與水分脅迫之間的互作效應對水稻葉面積影響顯著。
2.3 水分脅迫對水稻根系的影響
2.3.1 對根系活力的影響
從表2可見,不同栽培方式和水分脅迫程度對水稻的根系氧化活力影響極顯著,表現為C2>C1,平均提高了20.5%;W2>W3>W4>W1,水分脅迫處理較對照W1分別提高了36% ~74.9%(C2)和27.9% ~59.7%(C1),但隨著水分脅迫的加重,根系活力增強的就越少,且處理間差異顯著。
水分脅迫對水稻根系活躍吸收面積影響顯著,對總吸收面積有影響但不顯著,栽培方式對根系吸收面積影響不顯著(表2)。其中處理W2和W3對根系總吸收面積效果最好,處理W2、W3和W4的根系活躍吸收面積較W1均增加。在4種水分處理下,壟作栽培均比傳統栽培根系吸收面積大,且根系活躍吸收面積之間差異顯著。同時C因素(栽培方式)和W因素(水分處理)之間的互作效應對水稻根系活力及吸收面積影響顯著。
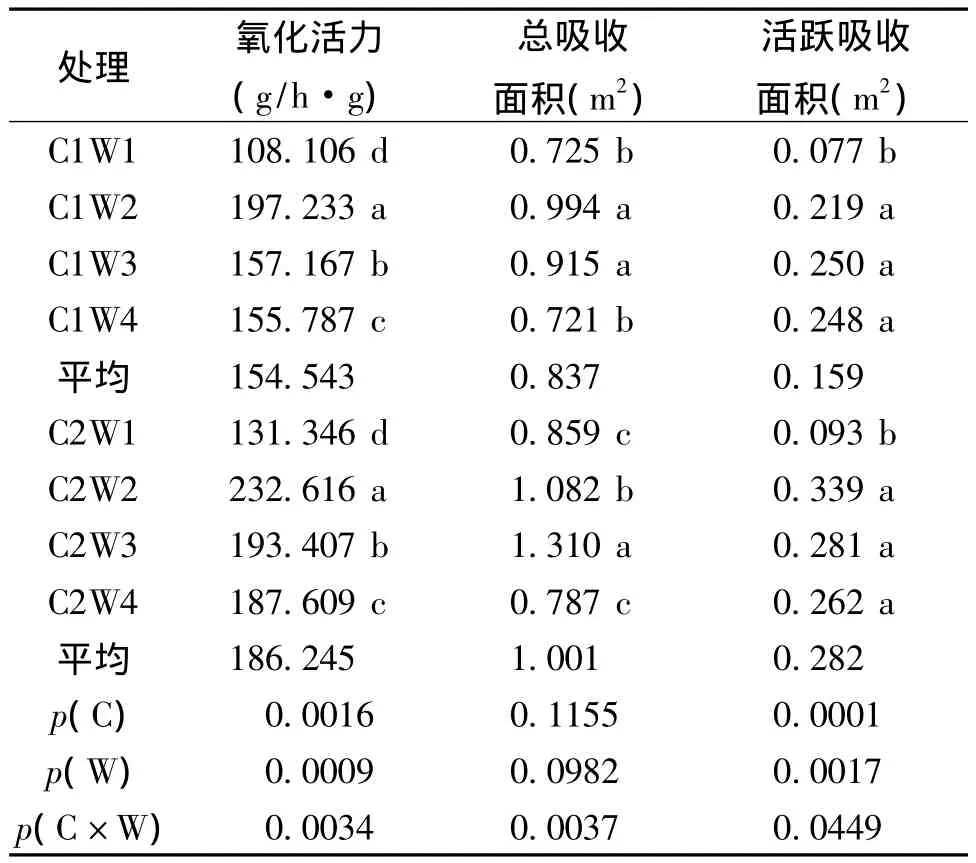
表2 不同水分脅迫下水稻根系活力及吸收面積比較Table 2 Comparison on activity and the absorption area of rice roots among different water stress treatments
2.3.2 對根系生長的影響
從表3可見,分蘗期水分脅迫對水稻總根數影響顯著,其中W2的總根數最多,W4最少,說明一定的水分脅迫有利于水稻根系的生長,但重度脅迫會削弱水稻根系生長。不同栽培方式對水稻總根數有影響,但差異不明顯,其中C2比C1平均增加了13.7%,說明壟作栽培較傳統栽培更有利于水稻根系的生長。
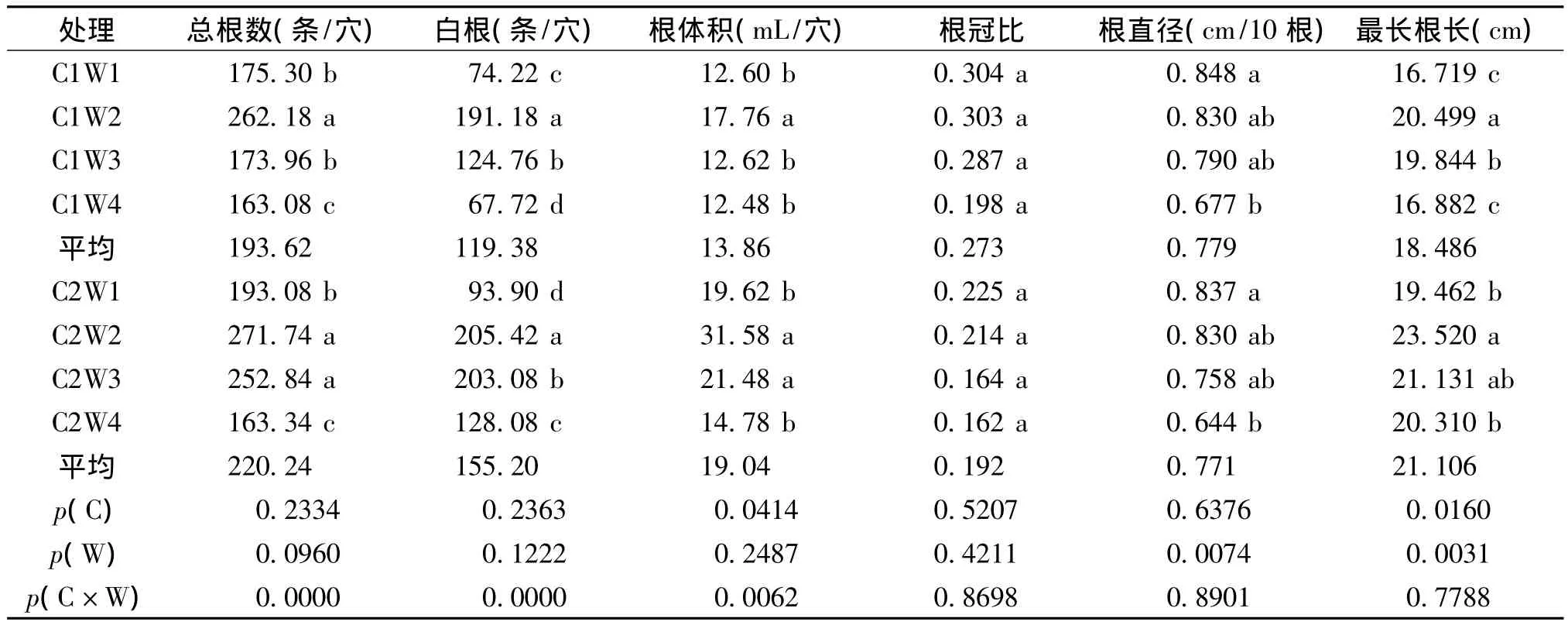
表3 不同水分脅迫下水稻的根系特征Table 3 Rice root characteristics under different water stress treatments
不同栽培方式及水分脅迫處理對水稻的白根數有影響但不顯著;不同栽培方式間也有差異,C2比C1的白根數平均增加了30.0%。說明一定的水分脅迫有利于延遲水稻根系的衰老,且壟作栽培比傳統栽培的效果更明顯。但水分脅迫過重,會加快水稻根系的衰老。
在不同水分脅迫處理下,水稻根體積變化為W2>W3>W1>W4,說明輕度、中度水分脅迫可以增加水稻根體積,重度脅迫則會降低根體積。兩種栽培方式下,C2比 C1的根體積平均增加了26.9%。
不同水分處理間的水稻根冠比為W1>W2>W3>W4;兩種栽培方式下,C2比C1的根冠比平均降低了29.7%。
不同水分處理間的水稻根直徑為W1>W2>W3>W4,說明水分脅迫能減小水稻的根直徑,且水分脅迫越重根直徑越小。兩種栽培方式下,C2比C1根直徑平均降低了1.0%。這是因為在逆境脅迫時細根是根系吸收的活性位點[12],它可以擴大根系與土壤的接觸面積來獲取更多的養分和水分[13~15]。而壟作使耕作層增厚,擴大了水稻根系的營養面積,有利于水稻向下生長[7],使根系直徑相對減小。
栽培方式和水分脅迫均對水稻最長根長影響顯著,表現為C2>C1,平均增加了14.2%;W2>W3>W4>W1,水分脅迫處理較對照分別提高了1.0% ~22.6%和4.4%~20.9%。
栽培方式與水分脅迫之間的互作效應對水稻總根數、白根數及根體積的影響顯著,對根冠比、根直徑及最長根長的影響則未達到顯著水平。
2.4 水分脅迫對水稻產量及水分利用效率的影響
表4可知,壟作栽培和水分脅迫對水稻的有效穗數影響顯著,表現為 C2>C1,W2>W1>W3>W4。其中有效穗數以W2最多,明顯高于W3和W4,說明輕度水分脅迫有利于提高水稻的有效穗數,而中度和重度水分脅迫則降低了水稻的有效穗數;實粒數也表現為W2>W1>W3>W4,說明分蘗期適度的水分脅迫對穎花分化沒有影響,相反可提高水稻的實粒數;水分脅迫對水稻的結實率有影響,輕度和中度脅迫提高了水稻的結實率,重度脅迫則會降低水稻的結實率;輕度的水分脅迫可提高水稻千粒重增加水稻產量,產量分別提高14.2%和16.1%,中度、重度脅迫則降低水稻產量,分別降低了23.5%和21.3%、41.6%和45.4%。分蘗期水分脅迫造成水稻產量低的原因主要是有效穗數和結實率降低所致。

表4 不同水分脅迫處理下的水稻產量及水分利用效率Table 4 Yield and water use efficiency of rice under different water stress treatments
不同的水分脅迫強度下水分利用效率以W2最高,顯著高于其它處理,這是因為W2的產量較W1、W3和W4高,W2耗水量低于W1處理10.4%和10.3%,W3的耗水量低于 W1處理 12.3%和11.4%,W4的耗水量低于 W1處理 15.6%和16.1%,但其水分的節省幅度低于產量下降的幅度,所以其水分利用效率較低。因此,輕度的水分脅迫可以提高水稻產量,提高水分利用率,達到節本增效的目的。
不同栽培方式下,壟作栽培比傳統栽培表現更好,其中產量平均提高15.2%,水分利用率平均提高17.3%,說明壟作栽培有利于提高水稻的產量及水分利用效率。
同時C因素(栽培方式)和W因素(水分處理)之間的互作效應對水稻產量和實粒數的影響達到顯著水平,對有效穗數、結實率、千粒重、耗水量及水分利用效率的影響則未達到顯著水平。
3 討論與結論
3.1 討論
分蘗期水分脅迫影響水稻分蘗數、葉齡和葉面積等生育指標,從而影響水稻產量性狀,影響的程度隨水分脅迫發生的階段、程度及持續時間而異。余叔文等[16]研究認為,分蘗后期和拔節期受重度干旱后沒有減產甚至有增產的趨勢;徐富賢等[17]認為,水稻分蘗期受干旱影響水稻產量的土壤含水量臨界值約為相對含水量的36%;陳國林[18]研究表明,適當的節水灌溉可使植株體內水分狀態得到調節,又能延緩下位葉的衰老,增加光合作用的時間,生育后期根系活力得到提高,協調了水稻高產與根系早衰的矛盾,生殖生長期干物質積累增加,有利于子粒的形成,最終獲得高產;水分脅迫對水稻分蘗的影響貫穿于整個生長期,適度的水分虧缺有利于水稻的分蘗,甚至結實率、千粒重等都較高,但過度的缺水則會抑制分蘗,影響有效分蘗數,從而導致減產。
壟作梯式栽培改變了土壤的微地形,可以改善土壤透氣性,促上控下的作用增強,使水稻根系發達,根系活力增強,吸收能力增強;擴大了葉片總受光面積,提高土壤溫度,促進有益微生物的活動,增加有效養分,土體水、肥、氣、熱協調,從而能提高水稻的抗逆性[8,19~21]。
3.2 結論
本試驗研究表明,分蘗期輕度水分脅迫有利于水稻的生長,能有效增加水稻的有效分蘗數、根系氧化活力、吸附面積,促進根系生長,中度水分脅迫對水稻的影響較常規灌溉有較小幅度的下降或者上升,但影響不顯著,而重度脅迫則影響顯著。分蘗期輕度水分脅迫對水稻的株高及葉面積指數影響不大,而中度和重度脅迫則嚴重制約著水稻株高的生長和葉面積的擴展。輕度水分脅迫可增加水稻的實粒數、千粒重及結實率,水稻產量提高14.2%和16.1%,耗水量降低10.4%和10.3%,水分利用率提高21.8%和22.7%,而中度水分脅迫則水稻產量降低23.5%和21.3%,耗水量降低12.3%和11.4%,水分利用率下降12.2%和11.8%,重度水分脅迫則水稻產量降低41.6%和45.4%,耗水量降低15.6%和16.1%,水分利用率下降30.4%和35.3%,產量下降幅度高于耗水節約幅度。兩種栽培方式下,壟作梯式栽培較傳統栽培水稻的千粒重、結實率、產量、耗水量及水分利用率要高,說明壟作栽培更有利于水稻的生長。因此,分蘗期輕度控水和采用壟作栽培有利于水稻生長和產量的提高。
[1]邵璽文,張瑞珍,齊春艷,等.拔節孕穗期水分脅迫對水稻生長發育及產量的影響[J].吉林農業大學學報,2004,26(3):237 -241.
[2]趙正宜,遲道才,劉中琦,等.水分脅迫對水稻生長發育影響的研究[J].沈陽農業大學學報,2000(2):214-217.
[3]王紹華,曹衛星,姜 東,等.水稻強化栽培對植株生理與群體發育的影響[J].中國水稻科學,2003,17(1):31-36.
[4]徐富賢,熊 洪,洪 松,等.水稻本田分蘗期受旱對其生育影響的研究[J].四川農業大學學報,2000,18(1):28-30.
[5]楊健昌,王 維,王志琴,等.水稻旱秧大田期需水特性與節水灌溉指標研究[J].中國農業科學,2000,33(2):34-42.
[6]黃文江,黃義德,王紀華,等.水稻旱作對其生長量和經濟產量的影響[J].干旱地區農業研究,2003,21(4):15-19.
[7]劉建松.水稻壟作栽培應用效果研究[J].中國農學通報,2002,18(6):16 -21.
[8]黃慶格.水稻壟作栽培的關鍵技術及其效應分析[J].廣西農業科學,1995(4):151 -152.
[9]劉剛才,張先婉,曾覺廷,等.川中丘陵區壟作土壤抗旱機理的研究[J].土壤通報,1997,28(6):248 -250,253.
[10]王昌全,魏成明,李廷強,等.不同免耕方式對作物產量和土壤理化性狀的影響[J].四川農業大學學報,2001,19(2):152 -154.
[11]張憲政,陳鳳玉,王榮富,等.植物生理學實驗技術[M].沈陽:遼寧科學技術出版社,1994.51-75.
[12]Zobel RW.Sensitivity analysis of computer based diameter measurement from digital images [J].Crop Sci,2003,43:583 -591.
[13]Huang B,Eissenstat DM.Lin king hydraulic conductivity to anatomy in plants that vary in specific root length[J].J Am Soc Hortic Sci,2000,125:260 -264.
[14]Benjamin JG,Nielsen DC.Water deficit effects on root distribution of soybean,field peaand chickpea[J].Field Crop Res,2006,97:248 -253.
[15]Peman J,Voltas J,GilPelegrin E.Morphological and functional variability in the root system of Quercusilex L.subject to confinement:consequences for afforestation[J].Ann For Sci,2006,63:425 -430.
[16]余叔文,陳景治,龔燦霞,等.不同生長期土壤干旱對水稻的影響[J].作物學報,1962(4):399-409.
[17]徐富賢,熊 洪,洪 松,等.水稻本田分蘗期受旱對其生育影響的研究[J].四川農業大學學報,2000,18(1):28-30.
[18]陳國林.水稻節水灌溉的生理生態效應研究[J].江西農業大學學報,1996,18(2):161-166.
[19]朱宏宇,高 波,張蘇萍,等.不同移栽方式對水稻產量及效益的影響[J].安徽農業科學,2007,35(29):9188-9190.
[20]王長清.冷浸爛泥田水稻壟作覆膜栽培試驗示范[J].湖北農業科學,1998(4):31 -34.
[21]章秀福,王丹英,屈衍艷,等.壟畦栽培水稻的植株形態與生理特性研究[J].作物學報,2005(6):742-748.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
礦山安全信息(2022年40期)2022-04-07 02:16:52
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代水產(2021年10期)2021-12-05 16:31:48
軍事文摘(2021年22期)2021-11-26 00:43:51
當代陜西(2021年2期)2021-03-29 07:41:24
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
聚氯乙烯(2018年9期)2018-02-18 01:11:34
