H7N9 亞型禽流感病毒非結構蛋白1(NS1)基因序列分析及2013上海分離株NS1的原核表達
2014-11-29 04:16:06婁文靜張道軍史云崔家駿吳志豪戚麗華靳連群劉雪林宋宏彬張傳福
生物技術通訊 2014年5期
婁文靜,張道軍,史云,崔家駿,吳志豪,戚麗華,靳連群,劉雪林,宋宏彬,張傳福
軍事醫學科學院 疾病預防控制所,北京 100071
H7N9亞型禽流感是甲型流感病毒中的一種,自暴發以來,在多個國家時有流行和散發,對養禽業造成了嚴重損失。2013 年我國出現首例人感染H7N9亞型禽流感病毒事件[1-2];截至目前,共報道143例確診病例,分布于北京、上海、江蘇、浙江等12 個省市,其中死亡45人,病死率高達31.5%[3]。因此,通過對今年暴發的人源H7N9 亞型禽流感病毒與禽源H7N9亞型病毒的基因同源性和系統進化分析,繼而研究監測并尋找對抗該病毒的方法是當務之急。
甲型流感病毒基因組是分為8 個節段的單股負鏈RNA,其中第8 段非結構蛋白基因是其基因組中最小的基因節段,編碼非結構蛋白NS1和NS2[4]。NS1 存在于病毒感染的細胞中,在正常病毒粒子中不存在,經滅活疫苗免疫的動物也不會產生。因此,針對NS1 的抗體可能只在流感病毒的宿主中存在,據此可以區分滅活疫苗免疫動物和野生型毒株感染動物[5]。此外,NS1 還具有抑制宿主蛋白合成、通過與宿主蛋白相互作用來增強病毒的致病性和毒力[6]、拮抗干擾素(IFN)產生及通過抑制絲氨酸/蘇氨酸蛋白激酶(PKR)-IFN 途徑和PI3K 激活途徑下調感染細胞凋亡[7]等功能。上述特點使NS1 在甲型流感病毒的檢測和抗病毒研究中有著廣闊的應用前景。
基于以上背景,我們克隆、表達了人源甲型流感病毒H7N9亞型NS1基因,為進一步研究其功能及基于NS1蛋白的抗病毒藥物奠定基礎。
1 材料與方法
1.1 材料
病毒株A/Shanghai/4664T/2013(H7N9)基因組cDNA 由本實驗室保存;大腸桿菌DH5α和BL21(DE3)、克隆載體pMD18-T、限制性內切酶、Taq酶、瓊脂糖凝膠回收試劑盒、T4DNA 連接酶、小量提取質粒試劑盒購自TaKaRa 公司;原核表達載體pET28a 購自Invitrogen 公司;DNA marker 購自Fermentas公司;其他試劑均為進口分裝或國產分析純。
1.2 NS1基因的擴增
根據GenBank 報道的A/Shanghai/4664T/2013(H7N9)株NS1基因序列設計PCR 引物,上游引物為5'-CCCATATGGGAGATATACCATGGGCA-3'(劃線部分為引入的NdeⅠ限制性酶切位點),下游引物為5'-GGAATTCCCGGAGCTCGAATTCTTA-3'(劃線部分為引入的EcoRⅠ限制性酶切位點)。擴增程序:94℃預變性30 s;94℃變性30 s,55℃退火1 min,72℃延伸2 min,30 個循環;72℃延伸5 min。將擴增產物行1%瓊脂糖凝膠電泳檢測。
1.3 目的基因的克隆
從瓊脂糖凝膠中回收PCR 產物,將純化的PCR產物片段與pMD18-T 克隆載體連接成重組質粒pMD18-T-NS1,轉化大腸桿菌DH5α感受態細菌,挑取白斑進行PCR鑒定,對陽性克隆進行序列測定,測序結果在NCBI網站在線比對分析。
1.4 重組表達質粒的構建與鑒定
將pMD18-T-NS1、pET28a 質粒分別用EcoRⅠ/NdeⅠ雙酶切,用T4DNA 連接酶將酶切后的NS1基因與pET28a載體連接,轉化大腸桿菌BL21(DE3)感受態細胞,構建重組表達載體pET28a-NS1,經酶切鑒定后,挑選陽性克隆質粒送中科西林生物有限公司進行基因序列測定,并進行序列分析。
1.5 NS1蛋白的誘導表達與表達產物檢測
挑取經鑒定的陽性單克隆于5 mL LB(Kan+)培養基中,37℃振蕩培養過夜,次日以1∶50 的比例轉種,擴大培養至菌液D600nm達0.5~0.7 時,加入終濃度為0.5 mmol/L 的IPTG,于37℃誘導表達2~3 h 后取菌液,12 000 r/min 離心5 min,棄上清,沉淀用PBS反復洗滌2 次,取1.5 g 菌體沉淀,用15 mL 菌體裂解液重懸,間歇超聲波冰浴破菌后,12 000 r/min離心 5 min,分別收集上清和沉淀,經12% SDSPAGE分析重組蛋白的表達形式,考馬斯亮藍染色。
1.6 Western印跡鑒定NS1蛋白
將SDS-PAGE 凝膠蛋白電轉移至PVDF 膜,經封閉液封閉后與1∶500 稀釋的His 單克隆抗體反應45 min,用含0.05% Tween-20 的PBS(PBST)洗滌3次,再與羊抗鼠IgG-HR 酶結合物室溫反應1 h,PBST充分洗滌后浸入底物顯色液中觀察顏色區帶。
2 結果
2.1 NS1基因的序列分析及系統進化樹分析
從GenBank 獲 得2006~2013 年不同 來源的H7N9 型病毒,包括2013 年在我國上海、安徽、江蘇、浙江、福建、廣東、臺灣等地暴發的H7N9 亞型禽流感病毒的NS1序列,用MEGA 5.0構建NJ進化樹(圖1)。從拓撲結構來看,A/emperor goose/Alaska/44063-061/2006(H7N9)與其余H7N9 型病毒分離株的NS1基因序列分為2 個進化分支。根據獲得的NS1基因核苷酸序列及推導的編碼氨基酸序列進行同源性比較,A/emperor goose/Alaska/44063-061/2006(H7N9)株與2013 年暴發的H7N9 型禽流感病毒NS1基因的核苷酸同源性僅為68.5%~70.6%,氨基酸序列同源性為51.1%~96.3%;與2007~2012 年暴發的H7N9 型禽流感病毒NS1基因的核苷酸同源性為70.5%~71.7%。
進化樹表明,2013 年的H7N9 亞型分離株都屬于基因群A 群,而2007~2012 年的分離株屬于基因群B 群。2013 年我國的H7N9 病毒分離株(A 群)其NS1基因存在以A/Wuxi/1/2013(H7N9)為代表和以A/Guangdong/1/2013(H7N9)為代表的2 個不同的亞分支;而2013 年中國大陸的H7N9 型禽源分離株其NS1基因都屬于A/Wuxi/1/2013(H7N9)為代表的分支,尚未發現屬于A/Guangdong/1/2013(H7N9)分支的2013 年禽源H7N9 病毒分離株。根據獲得的NS1基因核苷酸序列及推導出的氨基酸序列進行同源性比較,以A/Wuxi/1/2013(H7N9)為代表的分支的病毒分離株NS1核苷酸同源性為99.2%~100%,氨基酸同源性 為98.5%~100% ;以A/Guangdong/1/2013(H7N9)為代表的分支的病毒分離株NS1基因的核苷酸同源性為95.0%~96.6%,氨基酸同源性為94.3%~98%;同源性比較結果表明,2013 年的H7N9 型病毒分離株NS1基因高度保守。由B 群分離株的進化分支來看,2008 年的危地馬拉分離株、2009 年的江西分離株、2009 年的捷克分離株、2011 年的韓國分離株等屬于不同的進化分支,說明H7N9 型禽流感不僅具有時間相關性,而且具有地域相關性。
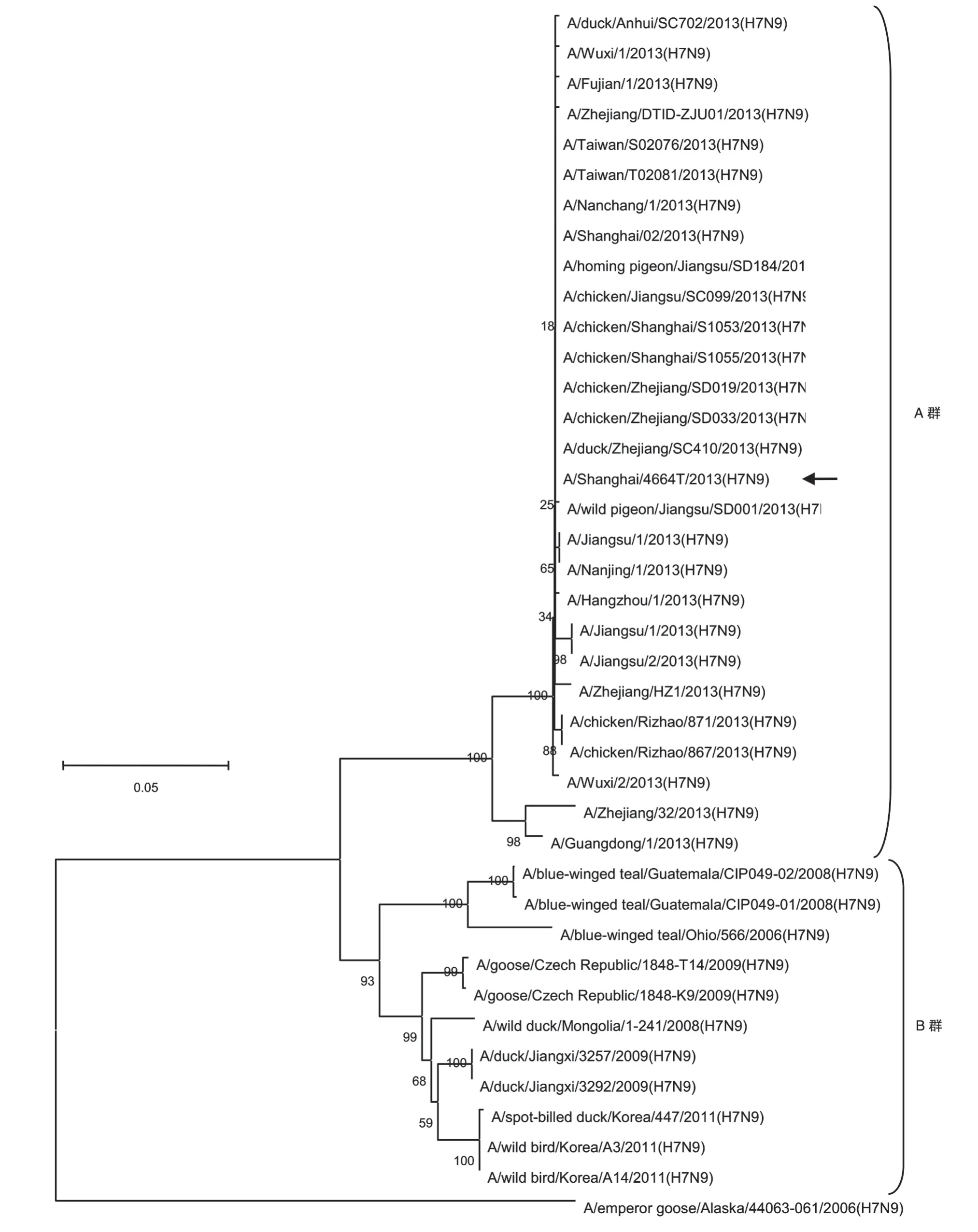
圖1 H7N9流感病毒NS1基因進化樹
2013 年暴發的H7N9 型禽流感病毒分離株(A群)與2007~2012 年暴發的H7N9 病毒分離株(B 群)屬于不同的進化分支,兩者間NS1基因的核苷酸同源性為86.4%~90.7%,氨基酸序列同源性為72%~87.3%。A 群與B 群在進化上的顯著差異暗示甲流病毒變異速度快這一特征,說明今年暴發的人源H7N9 型禽流感與之前暴發的禽源H7N9 型禽流感在進化上有顯著區別。因此,對同一種H7N9 型禽流感病毒的不同地域、親緣關系的研究,有助于進一步了解病毒的進化特征,為未來基于NS1 蛋白的抗病毒藥物及疫苗的研發奠定基礎。
2.2 NS1基因的擴增及克隆質粒鑒定
以A/Shanghai/4664T/2013(H7N9)基因組cDNA為模板,用設計的特異性引物擴增NS1基因全長,擴增片段與預期大小相符。重組克隆質粒pMD18-TNS1 轉化菌的PCR 產物經1%瓊脂糖凝膠電泳分析,在約680 bp 處可見特異性條帶,與預期相符。測序結果與A/Shanghai/4664T/2013(H7N9)株NS1基因序列一致。
2.3 重組表達質粒的鑒定
提取重組質粒pET28a-NS1,經限制性內切酶EcoRⅠ/NdeⅠ酶切后,瓊脂糖凝膠電泳得到2 個片段,其中小片段與目的基因片段大小相符(圖2),測序結果表明,NS1基因以正確的方式插入pET28a 載體中,且序列正確。
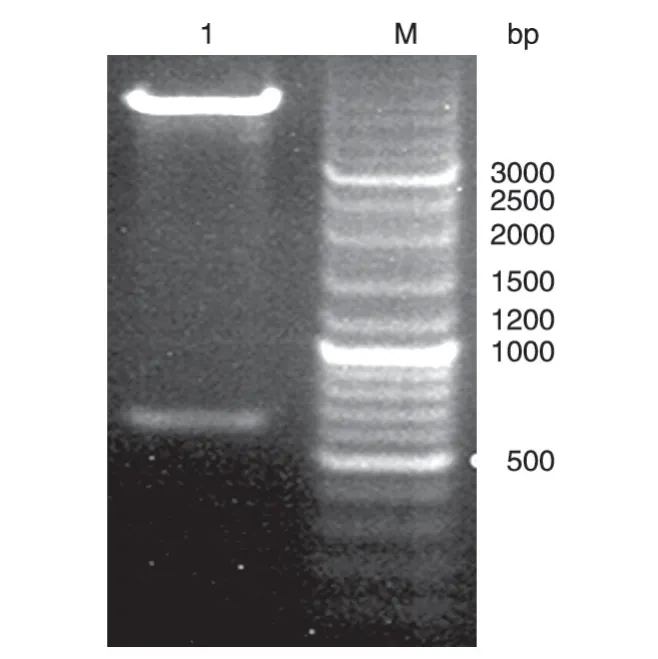
圖2 重組表達質粒的酶切產物電泳圖
2.4 NS1蛋白的誘導表達與SDS-PAGE分析
含pET28a-NS1 重組質粒的大腸桿菌BL21(DE3)經0.5 mmol/L IPTG 誘導后超聲波裂解,SDS-PAGE 分析(圖3)表明有相對分子質量為25×103的蛋白表達,與預期一致,而未經IPTG 誘導的對照樣本則沒有預期蛋白表達。從圖3可以看出,NS1融合蛋白在菌體裂解后的上清和沉淀中均有,但主要以包涵體形式存在。
2.5 表達蛋白的特異性鑒定
為了驗證相對分子質量為25×103的蛋白帶是否由NS1基因表達,用Western 印跡進行了表達產物的特異性檢測,結果如圖4,IPTG 誘導樣本有特異條帶,而未加IPTG 誘導的大腸桿菌未見特異條帶,證明該蛋白帶系NS1基因表達產物。
3 討論
自1988年在美國明尼蘇達州首次發現H7N9亞型禽流感以來[8],陸續在多個國家和地區有該病發生的報道,該型病毒在家禽和水禽中廣泛存在,已對養禽業造成了嚴重損失[9]。2013 年我國首次出現H7N9 亞型禽流感感染人的現象,截至目前已造成45人死亡[10]。2013 年10 月,浙江省又出現2例人感染禽流感病例。因此,對該病毒的研究應當予以重視。目前,相關研究多側重于H7N9 病毒血凝素,而對非結構蛋白及其基因的研究較為少見。非結構蛋白只在病毒感染的細胞中合成,并未包裝進病毒顆粒。在病毒感染早期,非結構蛋白就在病毒中大量表達且積聚在細胞核內,形成致密的晶體樣包涵體,在感染晚期的細胞漿內也有存在,刺激機體產生抗NS1 蛋白的抗體[11]。現有研究表明,NS1 蛋白在抑制宿主細胞蛋白質合成、增強病毒mRNA 翻譯的效率、誘導細胞凋亡和抑制干擾素等方面具有重要作用。因此,拮抗NS1蛋白的功能,對抑制甲型流感病毒的復制和限制病毒傳播具有良好的應用價值[12]。
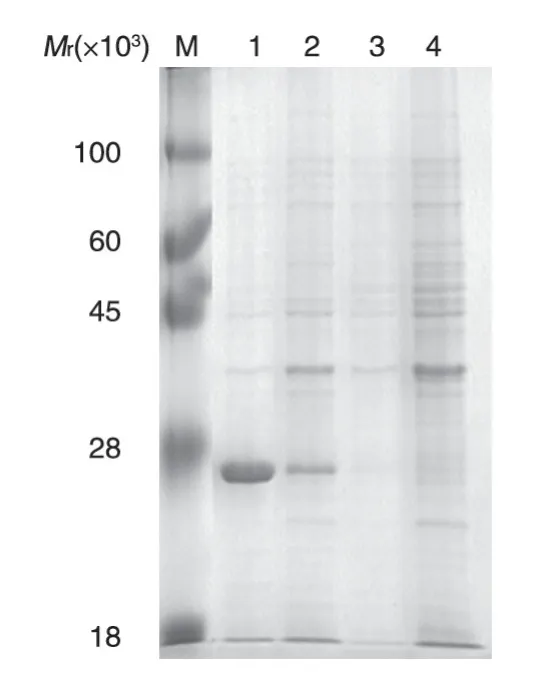
圖3 SDS-PAGE檢測pET28a-NS1在大腸桿菌BL21(DE3)中的誘導表達產物

圖4 表達產物的Western印跡
我們擴增了A/Shanghai/4664T/2013(H7N9)分離株的NS1 基因,并與從GenBank 下載的其他H7N9病毒分離株的NS1基因片段序列和蛋白序列做了對照分析,發現H7N9 流感病毒的NS1 基因分為A/emperor goose/Alaska/44063-061/2006(H7N9)獨立分支和基因群A、B 群,其中A、B 群兩群群內的核苷酸序列同源性分別為95%~100%和92.1%~100%,而兩群之間的核苷酸同源性僅為86.4%~90.7%。同源性比較表明,2013 年H7N9 禽流感病毒分離株同源性高達95%以上,在進化上高度保守,與之前的H7N9型病毒分離株有明顯的進化區別。
隨著天氣轉冷,人感染H7N9 禽流感病例可能會繼續出現。此外,目前禽間病毒傳播并未得到控制,且禽類感染H7N9 病毒后很難發現,難以在早期控制禽類疫情[13]。因此,對H7N9禽流感進行監測是預防與控制其暴發的一個重要環節。我們以此為出發點,擴增了人源H7N9 亞型禽流感病毒的NS1基因,對其進行基因克隆,在大腸桿菌BL21(DE3)中高效表達了NS1 融合蛋白,重組蛋白可以與流感病毒的NS1蛋白單克隆抗體反應。這一結果有助于研究H7N9亞型禽流感病毒非結構蛋白的活性、對疫苗免疫和野毒感染的動物診斷,并為以NS1 蛋白為基礎的抗病毒藥物的研制奠定了基礎,為將來有效地進行禽流感的監測工作邁出了關鍵一步。
[1]Parry J.H7N9 avian flu infects humans for the first time[J].Br Med J,2013,346:2151.
[2]Parry J.H7N9 avian flu kills seven and infects 23 in China[J].Br Med J,2013,346:2222.
[3]Shi Y,Zhang W,Wang F,et al.Structures and receptor binding of hemagglutinins from human-infecting H7N9 influenza viruses[J].Science,2013,342(6155):243-247.
[4]Skehel J J.Polypeptied synthesis in influenza virus-infected cells[J].Virology,1972,49:23-36.
[5]Ozaki H,Sugiura T,Sugita S,et al.Detection of antibodies to the nonstructural protein(NS1) of influenza A virus allows distinction between vaccinated and infected horses[J].Vet Microbiol,2001,82:111-119.
[6]Wolff T,O'Neill R E,Palese P.Interaction cloning of NS1-I,a human protein that binds to the nonstructural NS1 proteins of influenza A and B viruses[J].J Virol,1996,70(8):5363-5372.
[7]Zhirnov O P,Konakova T E,Wolff T,et al.NS1 protein of influenza A virus down-regulates apoptosis[J].J Virol,2002,76(4):1617-1625.
[8]Obenauer J C,Denson J,Mehta P K,et al.Large-scale sequence analysis of avian influenza isolates[J].Science,2006,311(5767):1576-1580.
[9]Nishiura H,Mizumoto K,Ejima K.How to interpret the transmissibility of novel influenza A(H7N9):an analysis of initial epidemiological data of human cases from China[J].Theor Biol Med Model,2013,10:30.
[10]Song P P,Xia J F,Gao J J,et al.Measures to combat H7N9 virus infection in China:Live poultry purchasing habits,poultry handling,and living conditions increase the risk of exposure to contaminated environments[J].Biosci Trends,2013,7(4):168-171.
[11]Nietzert E,Beck E,Ke Mello P A,et al.Expression of the aphthovirus RNA polymerase gene in Escherichia coli and tis use with other bioengineered nonstructural antigens in detection of late persistent infection[J].Virology,1991,1847:799-804.
[12]Suarez D L.Overview of avian influenza DIVA test strategies[J].Biologicals,2005,33(4):221-226.
[13]Parry J.H7N9 virus is more transmissible and harder to detect than H5N1,say experts[J].Br Med J,2013,346:2568.
