腸出血性大腸桿菌毒力因子Z1444的原核表達及其絲/蘇氨酸激酶活性驗證
2014-10-27 09:04:40李嶄李濤陳芳紅劉雄周育森王慧
生物技術(shù)通訊 2014年3期
李嶄,李濤,陳芳紅,劉雄,周育森,王慧
1.廣西醫(yī)科大學(xué),廣西 南寧 5300002;2.軍事醫(yī)學(xué)科學(xué)院 微生物流行病研究所,病原微生物生物安全國家重點實驗室,北京 100071
腸出血性大腸桿菌(enterohemorrhagic Esche?richia coli,EHEC)O157∶H7作為一種重要的食源性致病菌,其感染特點為世界流行性、高發(fā)病率與病死率,以及抗生素治療會加劇病情等[1]。自美國1982年首次暴發(fā)流行以來,許多國家相繼發(fā)生了EHEC O157∶H7的感染流行[2]。現(xiàn)今其感染已成為一個全球性的公共衛(wèi)生問題,受到各國廣泛關(guān)注[3]。對該菌致病機制的研究,已成為世界醫(yī)學(xué)界的關(guān)注點[4-6]。
EHEC O157∶H7的致病機制與其基因組中某些特殊區(qū)域有關(guān),這些區(qū)域可編碼與EHEC致病性有關(guān)的毒力因子,而毒力島就是這些特殊基因的編碼區(qū)[7]。O157∶H7毒力島包括編碼eae基因的LEE島、前噬菌體上編碼志賀樣毒素(Shiga-like toxin,SLT)的slt基因和大質(zhì)粒(pO157)編碼的hly、katP、espP、toxB、stcE基因等[8],目前對EHEC致病機制的研究主要集中在毒力島編碼系統(tǒng)上[9]。通過對毒力島的研究發(fā)現(xiàn)了一些重要的毒力因子,但迄今所發(fā)現(xiàn)的毒力因子不足以解釋0157∶H7的全部致病過程[10]。
我們通過對EHEC毒力島編碼序列的掃描,在毒力島前噬菌體編碼區(qū)志賀樣毒素基因slt上游位置發(fā)現(xiàn)了功能未知的Z1444蛋白基因[11-13]。通過毒力因子基因同源比對,分析致病基因保守區(qū)域,預(yù)測EHEC染色體一段完整基因ORF Z1444具有潛在的致病性,該基因編碼的蛋白很可能參與細菌與宿主的相互作用,并且可能是具有Ⅲ型分泌系統(tǒng)的致病菌重要的效應(yīng)蛋白(毒力因子)。因此,研究0157∶H7 Z1444蛋白,對于進—步了解細菌與宿主的相互作用具有重要意義。
1 材料與方法
1.1 材料
EHEC O157∶H7 EDL933株和 pET-28a(+)載體為本室保存;大腸桿菌DH5α、BL21(DE3)感受態(tài)細胞,pEASY-T1載體,Taq酶,dNTP及DNA marker購自TransGen公司;質(zhì)粒小提試劑盒和瓊脂糖凝膠回收試劑盒購自北京博邁德公司;NdeⅠ、XhoⅠ限制性內(nèi)切酶及T4DNA連接酶購自NEB公司;His抗體和磷酸絲氨酸/蘇氨酸抗體分別購自CST與Ab?cam公司。
1.2 Z1444基因的擴增
根據(jù) GenBank中的 Z1444基因序列(ID:959087)設(shè)計引物 Z1444(up)(5'-CATATGCTAACT CCATACAAAAG-3')和 Z1444(down)(5'-CTCGAG GTTATCCTTTAATAACCTATACTG-3'),分別在上下游引物中引入NdeⅠ、XhoⅠ酶切位點(下劃線序列)。以EHEC O157∶H7全菌裂解液為模板,Z1444(up)、Z1444(down)為引物擴增目的基因。PCR條件:95℃預(yù)變性5 min;94℃變性30 s,53℃復(fù)性30 s,72℃延伸90 s,30個循環(huán);72℃延伸10 min,4℃停止。取25 μL PCR產(chǎn)物于10 g/L瓊脂糖凝膠中電泳(116 V),20 min后在紫外燈下觀察結(jié)果并拍照。
膠回收PCR產(chǎn)物,與pEasy-T1載體連接,轉(zhuǎn)入大腸桿菌DH5α感受態(tài)細胞,在LB培養(yǎng)基中過夜培養(yǎng)(37℃、200 r/min),次日從轉(zhuǎn)化平板上挑取單克隆菌落,以 Z1444(up)、Z1444(down)為引物進行 PCR篩選鑒定陽性菌株,陽性克隆送生工生物工程有限公司測序。
1.3 表達菌株pET-28a(+)-Z1444/BL21的構(gòu)建
1.3.1 表達質(zhì)粒pET-28a(+)-Z1444的構(gòu)建 將測序正確的克隆質(zhì)粒pEasy-T1-Z1444用NdeⅠ、XhoⅠ雙酶切,得到目的片段Z1444,膠回收,用T4DNA連接酶連接經(jīng)同樣雙酶切處理的pET-28a(+)載體,連接產(chǎn)物轉(zhuǎn)化大腸桿菌DH5α感受態(tài)細胞,在LB培養(yǎng)基中過夜培養(yǎng)(37℃、200 r/min),次日從轉(zhuǎn)化平板上挑取單克隆菌落,以Z1444(up)、Z1444(down)為引物進行PCR初篩,NdeⅠ、XhoⅠ雙酶切復(fù)檢,鑒定陽性菌株,陽性克隆送生工生物工程有限公司測序。
1.3.2 表達菌株的構(gòu)建 將測序正確的重組質(zhì)粒pET-28a(+)-Z1444轉(zhuǎn)化大腸桿菌BL21(DE3)感受態(tài)細胞,在LB培養(yǎng)基中過夜培養(yǎng)(37℃、200 r/min),次日挑取單克隆,以Z1444(up)、Z1444(down)為引物進行PCR鑒定。
1.4 目的蛋白的表達與純化
1.4.1 蛋白表達 鑒定為陽性的單克隆菌株和轉(zhuǎn)入空載體的陰性對照菌接入新鮮LB培養(yǎng)基,37℃培養(yǎng)至對數(shù)生長期前期時(D600nm約為0.4),加入IPTG至終濃度為1 mmol/L,25℃、200 r/min條件下誘導(dǎo)6 h,5000 r/min離心20 min收集菌體,用1/5體積的PBS重懸菌體,400W功率超聲4個循環(huán)(工作3 s,間隔5 s,循環(huán)99次),破碎菌體細胞,5000 r/min離心15 min,分離上清、沉淀,各取20 μL樣品進行SDSPAGE(5%濃縮膠和15%分離膠;恒壓110 V,濃縮15 min;恒壓180 V,分離1 h)。
1.4.2 蛋白純化 超聲裂解后上清過鎳柱(His?Trap FF crude 5 mL)純化,用400 mmol/L咪唑洗脫液洗脫,取純化后蛋白20 μL進行SDS-PAGE。
1.5 純化蛋白的功能初步驗證
1.5.1 His標簽蛋白的Western印跡 收集純化的蛋白,進行15%SDS-PAGE鑒定,并電轉(zhuǎn)移到醋酸纖維素(PVDF)膜上,用3%BSA封閉(37℃、2 h),加入1∶1000稀釋的兔源抗His抗體(一抗),4℃過夜,再加入1∶1000稀釋的HRP酶標二抗(抗兔),室溫輕搖1 h,用TBST洗膜后,在BIO-RAD凝膠成像系統(tǒng)中化學(xué)發(fā)光顯影。
1.5.2 純化蛋白的體外酶活實驗 GenBank中Z1444的蛋白功能注釋為絲/蘇氨酸蛋白激酶,因此選取絲/蘇氨酸通用底物髓鞘堿性蛋白(myelin ba?sic protein,MBP)進行測定[14]。首先將 MBP溶于酶活實驗低滲緩沖液[20 mmol/L Hepes(pH7.4),150 mmol/L NaCl,10 mmol/L MgCl2]中,然后加入Z1444蛋白,補加50 μmol/L ATP,于30℃反應(yīng)30 min后收集產(chǎn)物,進行15%SDS-PAGE后電轉(zhuǎn)移至PVDF膜上,用3%BSA封閉(37℃、2 h),加入1∶1000稀釋的鼠源磷酸絲氨酸/蘇氨酸抗體(一抗),4℃過夜,再加入1∶1000稀釋的HRP酶標二抗(抗鼠),室溫輕搖1 h,用TBST洗膜后,在BIO-RAD凝膠成像系統(tǒng)中化學(xué)發(fā)光顯影。
2 結(jié)果
2.1 pET-28a(+)-Z1444原核表達質(zhì)粒的構(gòu)建與鑒定
以EHEC O157∶H7全菌裂解液為模板,Z1444(up)、Z1444(down)為引物PCR擴增Z1444基因,經(jīng)瓊脂糖凝膠電泳,獲得約1047 bp的DNA片段,與預(yù)期一致(圖1)。將pET-28a(+)載體與PCR鑒定陽性的質(zhì)粒pEasy-T1-Z1444同時經(jīng)NdeⅠ/XhoⅠ雙酶切,各自膠回收產(chǎn)物用T4DNA連接酶連接,轉(zhuǎn)入大腸桿菌DH5α,涂板后挑取克隆酶切鑒定,可切出2條片段,分別為長度約5000 bp的空載體pET-28a(+)和1047 bp的Z1444條帶,重組質(zhì)粒pET-28a(+)-Z1444作為雙酶切前對照,符合預(yù)期結(jié)果(圖2)。完成后質(zhì)粒送生工生物工程有限公司測序,測序結(jié)果與NCBI數(shù)據(jù)庫收錄的相應(yīng)堿基序列的一致率為100%(序列略)。
2.2 重組Z1444的表達和純化
重組菌pET-28a(+)-Z1444/BL21經(jīng)IPTG誘導(dǎo)表達后制備全菌裂解液,分離沉淀和上清,取樣品進行SDS-PAGE鑒定,可見上清中在相對分子質(zhì)量約38×103處有明顯的蛋白條帶,與預(yù)期相符(圖3),而沉淀中對應(yīng)位置并無顯著條帶。轉(zhuǎn)入空載體的菌體裂解液作為陰性對照,經(jīng)相同處理后,對應(yīng)位置無目的條帶。因此,可以判斷獲得了原核表達的目的蛋白,且以可溶性形式存在。將重組菌全菌裂解液上清經(jīng)鎳柱純化,去除大量雜蛋白,得到較純的蛋白條帶(圖4),所得蛋白溶液經(jīng)反復(fù)凍融后沒有出現(xiàn)沉淀,可供后續(xù)試驗使用。
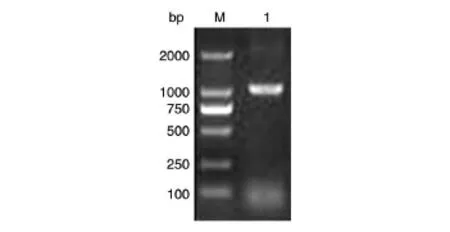
圖1 PCR擴增Z1444基因
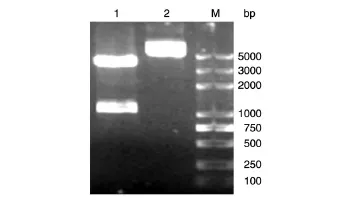
圖2 重組質(zhì)粒pET-28a(+)-Z1444的酶切鑒定
2.3 重組Z1444的絲/蘇氨酸激酶活性初步驗證
將純化的Z1444蛋白進行Western印跡,檢測His-Z1444蛋白表達,結(jié)果見圖5。同時將絲/蘇氨酸激酶通用磷酸化底物MBP與Z1444蛋白混合后補加ATP,30℃孵育30 min,完成酶促反應(yīng),用磷酸絲/蘇氨酸抗體進行Western印跡檢測,可見Z1444蛋白不僅可以磷酸化通用底物MBP,且Z1444蛋白本身發(fā)生了自磷酸化現(xiàn)象(圖6)。
3 討論
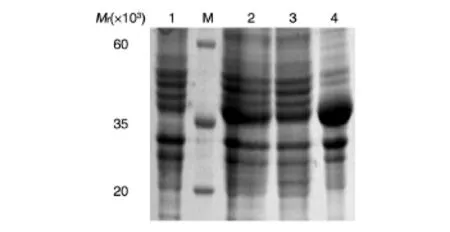
圖3 重組菌pET-28a(+)-Z1444/BL21的蛋白表達
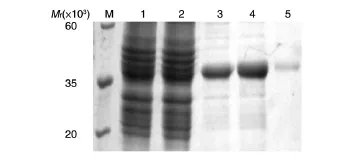
圖4 重組Z1444蛋白的純化

圖5 Western印跡檢測His-Z1444蛋白的表達

圖6 純化的Z1444蛋白的絲/蘇氨酸激酶活性驗證
我們通過寡核苷酸互補原理雙向設(shè)計引物,從大腸桿菌O157∶H7 EDLL933全基因組中釣取出Z1444基因,利用PCR擴增目的基因,選用帶有His標簽的pET載體系統(tǒng)構(gòu)建了重組表達質(zhì)粒,然后以大腸桿菌BL21(DE3)作為宿主菌進行蛋白表達。我們希望得到高水平表達的重組蛋白,且最好為可溶性形式,以利于蛋白純化及后續(xù)活性驗證。為此,我們進行了表達條件的優(yōu)化。在較高溫度如常規(guī)37℃誘導(dǎo)表達時發(fā)現(xiàn)重組蛋白大多形成包涵體,于是進行了溫度梯度誘導(dǎo)實驗,發(fā)現(xiàn)25℃誘導(dǎo)時重組蛋白為可溶性表達,產(chǎn)物表達量占上清總蛋白的40%左右。后續(xù)采用親和層析NTA His結(jié)合樹脂純化蛋白,配制6種濃度的咪唑洗脫液進行蛋白梯度洗脫純化,發(fā)現(xiàn)400 mmol/L咪唑洗脫液具有優(yōu)勢,可除去大部分雜蛋白,特異性較好,可獲得純度較高的蛋白。Z1444蛋白在原核系統(tǒng)中的高效可溶性表達,為后續(xù)蛋白活性研究提供了良好的實驗材料。
O157∶H7是EHEC的主要血清型,能引起人的出血性結(jié)腸炎和溶血性尿路綜合征[15]。EHEC可以產(chǎn)生毒力極強的志賀毒素,且具有黏附上皮細胞的能力,感染人或動物時黏附到腸黏膜上皮細胞,然后在此定居,釋放外毒素,從而引起疾病[16-17]。毒力島是與細菌致病性和毒力因子密切相關(guān)的基因簇[18-19],獲得EHEC O157:H7毒力島上某些基因的可溶性原核表達,為研究該基因的功能奠定了基礎(chǔ),同時有助于探索基因與菌株致病性之間的相互關(guān)聯(lián)。
綜上所述,我們構(gòu)建了pET-28a(+)-Z1444重組表達載體,并篩選獲得了高效原核表達工程菌,建立了簡便可行的蛋白純化工藝,獲得了濃度純度在90%以上的Z1444蛋白,為進一步研究Z1444蛋白在大腸桿菌與宿主相互作用時所起的功能奠定了基礎(chǔ)。我們還通過Western印跡驗證了GenBank對Z1444蛋白的功能注釋,發(fā)現(xiàn)該蛋白不僅可以磷酸化通用底物MBP,而且蛋白本身也發(fā)生自磷酸化現(xiàn)象。考慮到這些特殊的酶活效應(yīng),對其蛋白生物學(xué)活性的進一步驗證顯得尤為重要,這將有助于了解功能未知毒力因子在細菌感染時的作用機制。
[1]Orskov F,Orskov I.Escherichia coli serotyping and disease in man and animals[J].Can J Microbiol,1992,38(7):699-704.
[2]烏全明.腸出血性大腸桿菌O157感染防治研究進展[J].微生物學(xué)雜志,2004,5:96-98.
[3]Hacker J,Kaper J B.Pathogenicity islands and the evolution of microbe[J].Annu Rev Microbiol,2000,54:614-719.
[4]陳素良,郭逸秀,李勝奎,等.O157感染死亡病例的流行病學(xué)調(diào)查報告[J].現(xiàn)代預(yù)防醫(yī)學(xué),2001,28:169-174.
[5]Hedberg C.Food-related illness and death in the United States[J].Emerg Infect Dis,1999,5(6):840-842.
[6]McDaniel T K,Jarvis K G,Donnenberg M S,et al.A genet?ic locus of enterocyte effacement conserved among diverse en?terobacterial pathogens[J].Proc Natl Acad Sci USA,1995,92:1664-1668.
[7]楊琴,閻有功,曹軍皓.腸出血性大腸桿菌三型分泌系統(tǒng)及轉(zhuǎn)位效應(yīng)器蛋白的研究進展[J].預(yù)防醫(yī)學(xué),2008,12(2):112-233.
[8]周勇,萬成松.大腸桿菌O157:H7的毒力島與毒力因子的研究進展[J].微生物學(xué)免疫學(xué)進展,2006,34(2):58-62.
[9]Hacker J,Blum-Oehler G,Mühldorfer I,et al.Pathogenicity islands of virulent bacteria:structure,function and impact on microbial evolution[J].Mol Microbiol,1997,23:1089-1097.
[10]Johansen B K,Wasteson Y,Granum P E,et al.Mosaic struc?ture of Shiga-toxin-2-encoding phages isolated from Esche?richia coliO157:H7 indicatesfrequentgene exchange be?tween lambdoid phage genomes[J].Microbiology,2001,147:1929-1936.
[11]Campbell A,Botstein D.Evolution of the lambdoid phages[M]//Hendrix R W,Roberts J W,Stahl F W.Lambda II:vol II.New York:Cold Spring Harbor,1983:365-380.
[12]Datz M,Janetzki-Mittmann C,Franke S,et al.Analysis of the enterohemorrhagic Escherichia coli O157 DNA region con?taining lambdoid phage gene p and Shiga-like toxin structur?al genes[J].Appl Environ Microbiol,1996,62(3):791-797.
[13]Plunkett G 3rd,Rose D J,Durfee T J,et al.Sequence of Shiga toxin 2 phage 933W from Escherichia coli O157:H7:Shiga toxin as a phage late-gene product[J].J Bacteriol,1999,181(6):1767-1778.
[14]Hanks S K,Quinn A M,Hunter T,et al.The protein kinase family:conserved features and deduced phylogeny of the cata?lytic domains[J].Science,1988,241(4861):42-52.
[15]Karmali M A,Steele B T,Petric M,et al.Sporadic cases of haemolytic-uraemic syndrome associated with faecal cytotoxin and cytotoxin-producing Escherichia coli in stools[J].Lancet,1983,1(8325):619-620.
[16]蔣小平,鄧思敏,鐘巍,等.大腸埃希菌O157:H7致病因素的研究進展[J].病原生物學(xué),2012,40(11):6531-6533.
[17]王斌,葉冬青.耶爾森菌強毒力島[J].微生物學(xué)通報,2003,3(30):99-103.
[18]徐建國.病原性細菌的III型分泌系統(tǒng)、毒力島和基因組學(xué)[J].國外醫(yī)學(xué)微生物學(xué)分冊,1999,2(19):169-172.
[19]Elliott S J,Wainwright L A,McDaniel T K,et al.The com?plete sequence ofthe locusforenterocyte effacement(LEE)from enteropathogenic Escherichia coli E2348/69[J].Mol Micro?biol,1998,28:1-4.
