利用優(yōu)化的5′-RACE實(shí)驗(yàn)定位副溶血弧菌基因的轉(zhuǎn)錄起始位點(diǎn)
2014-10-27 09:04:40李杰張義全高鶴王麗胡小許周冬生
生物技術(shù)通訊 2014年3期
關(guān)鍵詞:實(shí)驗(yàn)
李杰,張義全,高鶴,王麗,胡小許 ,周冬生
1.中國疾病預(yù)防控制中心 傳染病預(yù)防控制所,傳染病預(yù)防控制國家重點(diǎn)實(shí)驗(yàn)室,北京 102206;2.軍事醫(yī)學(xué)科學(xué)院 微生物流行病研究所,病原微生物生物安全國家重點(diǎn)實(shí)驗(yàn)室,北京 100071
基因是遺傳物質(zhì)的最小功能單位,它通過轉(zhuǎn)錄和翻譯而對生物體表型產(chǎn)生專一性效應(yīng)。轉(zhuǎn)錄是DNA指導(dǎo)下的RNA合成過程,起始于DNA模板的一個(gè)特定起點(diǎn),即基因的轉(zhuǎn)錄起始位點(diǎn)。對原核生物而言,其mRNA 5′端的堿基即為其轉(zhuǎn)錄起始位點(diǎn)。
在前期研究中[1-5],我們通常采用引物延伸實(shí)驗(yàn)來確定基因的轉(zhuǎn)錄起始位點(diǎn)。但引物延伸實(shí)驗(yàn)需要利用放射性核素對特異性引物的5′端進(jìn)行標(biāo)記,這不僅極易產(chǎn)生放射性廢物污染,而且對實(shí)驗(yàn)者的健康也有較大傷害。因此,該方法并不是尋找基因轉(zhuǎn)錄起始位點(diǎn)的最優(yōu)選擇。5′-cDNA末端快速擴(kuò)增(5′-rapid-amplification of cDNA ends,5′-RACE)實(shí)驗(yàn)也可以用來確定基因的轉(zhuǎn)錄起始位點(diǎn),該方法通過在mRNA的5′端連接接頭(Adapter),而后聯(lián)合利用逆轉(zhuǎn)錄、巢式PCR和DNA測序技術(shù),即可最終確定基因的轉(zhuǎn)錄起始位點(diǎn),具有靈敏度高和特異性好的優(yōu)點(diǎn)。然而,目前市售5′-RACE試劑盒均是針對真核生物mRNA的帽子結(jié)構(gòu)而設(shè)計(jì)的。由于細(xì)菌的mRNA無帽子結(jié)構(gòu),市售試劑盒并不能直接應(yīng)用于原核生物。
副溶血性弧菌(Vibrio parahaemolyticus,VP菌)是一種廣泛分布于淺海海域及海產(chǎn)品中的革蘭陰性弧菌,也是引起食物中毒的首要病原菌,能引起以腹瀉為主要癥狀的急性胃腸炎[6]。VP菌具有多種毒力因子,主要包括Ⅲ型分泌系統(tǒng)(T3SS1和T3SS2)、直接耐熱溶血素(TDH)和TDH相關(guān)溶血素(TRH)。ToxR(VP0820編碼)是VP菌的毒力轉(zhuǎn)錄調(diào)控子,能調(diào)節(jié)TDH和T3SS1相關(guān)基因的表達(dá)[7-8]。VP菌具有很強(qiáng)的生物膜形成能力,以抵抗不利的生存環(huán)境。作為第二信使,c-di-GMP能促進(jìn)生物膜的形成。VP菌有數(shù)十個(gè)蛋白參與c-di-GMP的分子代謝,包括ScrC[9]、ScrG[10]及 VPA0198。ScrC 由操縱子 scrABC編碼,主要具有磷酸二酯酶活性,能降解c-di-GMP,抑制胞外多糖CPS(cps操縱子編碼,cpsA是其首基因)的合成,從而抑制生物膜的形成[9,11];ScrG也主要表現(xiàn)為磷酸二酯酶活性,參與降解c-di-GMP[10];VPA0198在c-di-GMP代謝中的功能還不是很清楚,可能主要參與c-di-GMP的合成。VP菌Ⅵ型分泌系統(tǒng)2(T6SS2)基因位點(diǎn)包含3個(gè)操縱子,分別為VPA1027-1024、VPA1043-1028和VPA1044-1046,該系統(tǒng)是其主要的黏附因子之一[12]。
在本研究中,我們以VP菌VP0820、VPA1027、scrC、scrG、cpsA及VPA0198為研究對象,在Ambion公司FirstChoice RLM-RACE試劑盒基礎(chǔ)上,通過優(yōu)化實(shí)驗(yàn)條件,摸索出一個(gè)適用于原核生物的5′-RACE實(shí)驗(yàn)方法。
1 材料和方法
1.1 材料
VP 菌 RIMD2210633(tdh+,KP+)和大腸桿菌DH5α由本實(shí)驗(yàn)室保存;HI肉湯培養(yǎng)基(2.5%Difco Heart Infusion Broth)購自BD公司;Ex Taq DNA聚合酶、dNTP、DNA marker為TaKaRa公司產(chǎn)品;PCR產(chǎn)物回收試劑盒為QIAGEN產(chǎn)品;DNA-free試劑盒和FirstChoice RLM-RACE試劑盒為Ambion公司產(chǎn)品;TRIzol試劑為Invitrogen公司產(chǎn)品;pGEM-T Easy Vector SystemⅠ(T載體)、Primer Extension System、fmol DNA Cycle Sequencing System等為Promega公司產(chǎn)品。
1.2 細(xì)菌培養(yǎng)與RNA提取
取20 μL甘油菌種接種于15 mL HI肉湯培養(yǎng)基中,37℃、200 r/min培養(yǎng)12~14 h(至平臺期),而后以1∶1000稀釋,接種至新鮮的15 mL HI肉湯中,37℃、200 r/min培養(yǎng)至D600nm約為1.0,收集菌體,用TRIzol試劑[1]提取細(xì)菌總RNA,總RNA質(zhì)量用1.5%甲醛變性瓊脂糖凝膠電泳監(jiān)測。
1.3 消化DNA
用DNA-free試劑盒去除總RNA中的DNA,50 μL 實(shí)驗(yàn)體系包括 5 μL 10×DNaseⅠ緩沖液、2.5 μL rDNaseⅠ、6~8 μg總RNA、1 μL RNase抑制劑,用無核酸酶水補(bǔ)足。37℃孵育30 min后,加入100 μL無核酸酶水和150 μL酚∶氯仿(1∶1),充分混勻,室溫12 000 r/min離心5 min,將上層水相轉(zhuǎn)入新的離心管中,加入150 μL氯仿,充分混勻,離心后取上層水相,加入150 μL異丙醇,充分混勻,12 000 r/min離心20 min,RNA沉淀用0.5 mL 70%乙醇漂洗1次,最后用10 μL無核酸酶水溶解。
1.4 Adapter的連接
將已知序列的寡核苷酸片段(Adapter)連接至RNA的5′端,反應(yīng)體系包括4 μL總RNA、1 μL 5′-RACE Adapter、1 μL 10×RNA 連接酶緩沖液、2 μL T4RNA連接酶(2.5 U/μL),補(bǔ)加無核酸酶水至10 μL。溫和混勻,37℃孵育1 h后,直接用于逆轉(zhuǎn)錄反應(yīng)。
1.5 逆轉(zhuǎn)錄反應(yīng)
將連接有Adapter的總RNA逆轉(zhuǎn)錄成cDNA,反應(yīng)體系包括 2 μL 上述經(jīng)處理的總 RNA、4 μL dNTP、2 μL 隨機(jī)十聚體(random decamers)、2 μL 10×逆轉(zhuǎn)錄緩沖液、1 μL RNase抑制劑、1 μL MMLV逆轉(zhuǎn)錄酶,補(bǔ)加無核酸酶水至20 μL。輕輕混勻,42℃孵育1 h。cDNA產(chǎn)物可直接用于PCR反應(yīng)。此步應(yīng)有不加逆轉(zhuǎn)錄酶的陰性對照。
1.6 巢式PCR反應(yīng)
采用巢式PCR方法擴(kuò)增目的基因。第一輪PCR體系包括0.5 μL cDNA模板、2.5 μL 10×PCR緩沖液、2 μL dNTP、10 μmol/L基因特異性外引物和位于Adpater上的外引物(表1)各1 μL、0.75 U DNA聚合酶,補(bǔ)加去離子水至25 μL。第二輪PCR體系包括0.5 μL第一輪PCR產(chǎn)物(無須純化回收)、5 μL 10×PCR緩沖液、4 μL dNTP、10 μmol/L基因特異性內(nèi)引物和位于Adpater上的內(nèi)引物(表1)各2 μL、1.25 U DNA聚合酶,補(bǔ)加去離子水至50 μL。PCR 反應(yīng)程序均為:95℃ 5 min;94℃ 30 s,60℃50 s,72℃ 50 s,30個(gè)循環(huán);72℃ 5 min。第二輪產(chǎn)物經(jīng)試劑盒回收后待用。
1.7 PCR產(chǎn)物連接至T載體
將回收的第二輪PCR產(chǎn)物直接連接至T載體,連接體系包括 100~200 ng DNA 片段、1 μL T載體、5 μL 2×DNA連接酶緩沖液、1 μL T4DNA連接酶(2.5 U/μL),補(bǔ)加去離子水至10 μL。輕微混勻后,置10℃連接4 h以上。采用熱沖擊法將連接產(chǎn)物轉(zhuǎn)入大腸桿菌DH5α,并用氨芐西林(100 μg/mL)抗性平板篩選抗性克隆,PCR鑒定(鑒定引物對同第二輪PCR)陽性者測序。
1.8 引物延伸實(shí)驗(yàn)
將與VPA1027 mRNA互補(bǔ)的特異性引物(表1)的5′端用[γ-32P]ATP(5000 Ci/mmol)進(jìn)行放射性標(biāo)記[13],并將其退火到mRNA上,進(jìn)而將其逆轉(zhuǎn)錄成cDNA。逆轉(zhuǎn)錄產(chǎn)物配伍測序條帶進(jìn)行6%聚丙烯酰胺凝膠電泳,放射自顯影后,通過引物延伸條帶的位置即可確定VPA1027轉(zhuǎn)錄起始位點(diǎn)。
2 結(jié)果
2.1 cDNA中無基因組DNA污染
總RNA用50 μL無核酸酶水溶解后,其濃度應(yīng)在1000 ng/μL以上,且經(jīng)瓊脂糖凝膠電泳檢測無降解后,方可滿足下一步實(shí)驗(yàn)需要。總RNA經(jīng)DNA-free試劑盒消化及連接Adapter后,直接逆轉(zhuǎn)錄成cDNA,而后通過RT-PCR方法驗(yàn)證cDNA中是否有DNA污染,結(jié)果見圖1,以cDNA+和基因組DNA為模板時(shí),均能擴(kuò)增出同理論值大小一致的產(chǎn)物(231 bp),而以cDNA-和水為模板時(shí),均不能擴(kuò)增出條帶,這說明cDNA中無基因組DNA污染,可用作5′-RACE實(shí)驗(yàn)的模板。

表1 本研究所用引物序列
2.2 靶基因的轉(zhuǎn)錄起始位點(diǎn)
采用巢式PCR擴(kuò)增目的基因片段,如圖2A所示,VPA1027、scrA、scrG、cpsA和VPA0198均擴(kuò)增出明亮的產(chǎn)物條帶,但scrA條帶拖尾現(xiàn)象較嚴(yán)重。將上述PCR產(chǎn)物直接與T載體連接,篩選鑒定陽性者送公司測序。圖2B~F分別為VPA1027、scrG、scrA、cpsA和VPA0198的測序結(jié)果,分別與標(biāo)準(zhǔn)序列(DNA)比對,并根據(jù)Adapter序列,可以得出它們的轉(zhuǎn)錄起始位點(diǎn)分別為G(-103)、G(-70)、T(-205)、C(-129)和G(-238)(翻譯起始位點(diǎn)為+1)。
2.3 引物延伸驗(yàn)證VPA1027的轉(zhuǎn)錄起始位點(diǎn)
圖3為VPA1027的引物延伸實(shí)驗(yàn)結(jié)果,可以看出,若以翻譯起始位點(diǎn)為+1,則VPA1027的轉(zhuǎn)錄起始位點(diǎn)為-103位的G,這與5′-RACE的結(jié)果一致,說明5′-RACE得到的轉(zhuǎn)錄起始位點(diǎn)是可靠的。
2.4 靶基因的啟動子區(qū)序列
通過轉(zhuǎn)錄起始位點(diǎn)查詢基因的核心啟動子區(qū),可以反向驗(yàn)證轉(zhuǎn)錄起始位點(diǎn)的正確性。圖4為VPA1027、scrG、scrA、cpsA和VPA0198的啟動子區(qū)序列。可以看出,scrG、cpsA和VPA0198均具有比較保守的-10(TATAAT)和-35(TTGACA)序列,這說明了其轉(zhuǎn)錄起始位點(diǎn)的可靠性;但VPA1027和scrA只有相對保守的-10序列,而-35序列保守性較差,這說明在生理?xiàng)l件下,它們的轉(zhuǎn)錄啟動可能需要反式激活因子的輔助才能完成。
3 討論
市售5′-RACE試劑盒是依據(jù)真核生物mRNA的帽子結(jié)構(gòu)而設(shè)計(jì)的。無帽子結(jié)構(gòu)的DNA片段、rRNA及tRNA的5′端磷酸基團(tuán)極易被牛小腸堿性磷酸酶(calf intestine alkaline phosphatase,CIP)水解,而帶帽子結(jié)構(gòu)的mRNA則不受影響。但是,mRNA的帽子結(jié)構(gòu)能被煙草酸焦磷酸酶(tobacco acid py?rophosphatase,TAP)水解,并留下5′端磷酸基團(tuán)。因此,可以依次用CIP和TAP處理總RNA,再利用T4RNA連接酶將一個(gè)已知序列的寡核苷酸序列(Adapter)連接至mRNA的5′端磷酸基團(tuán)上,并將其逆轉(zhuǎn)錄成cDNA,最后通過巢式PCR和DNA測序的方法即可確定mRNA的5′末端。由于原核生物的mRNA的無帽子結(jié)構(gòu),市售5′-RACE試劑盒并不能直接應(yīng)用于原核生物mRNA的5′末端堿基的確定。我們在Ambion公司FirstChoice RLM-RACE試劑盒的基礎(chǔ)上,拋棄CIP和TAP處理總RNA的步驟,利用Ambion公司的DNA-free試劑盒去除總RNA中污染的DNA,而后直接將Adapter接頭連接至mRNA 5′末端,再將總RNA逆轉(zhuǎn)錄成cDNA,最后通過巢式PCR及測序的方法,成功定位了副溶血弧菌VPA1027、scrA、scrG、cpsA和VPA0198的轉(zhuǎn)錄起始位點(diǎn),并通過引物延伸的方法驗(yàn)證了VPA1027的轉(zhuǎn)錄起始位點(diǎn)。優(yōu)化后的方案不僅操作更簡單,而且應(yīng)可適用于所有原核生物基因的轉(zhuǎn)錄起始位點(diǎn)定位。
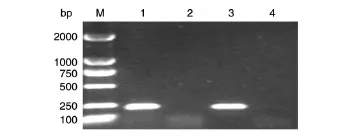
圖1 VP0820的RT-PCR產(chǎn)物1%瓊脂糖凝膠電泳圖
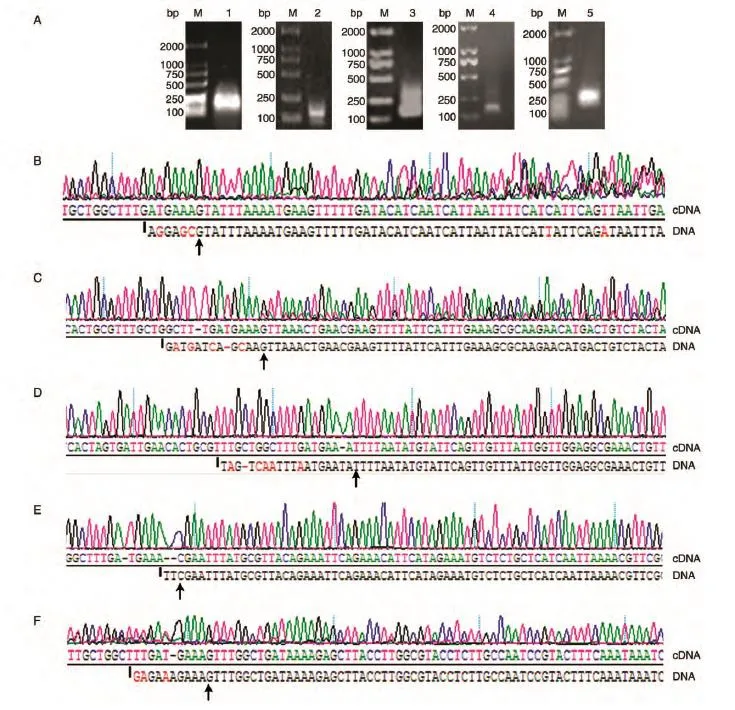
圖2 第二輪PCR產(chǎn)物1%瓊脂糖凝膠電泳圖(A)及測序圖譜(B~F)
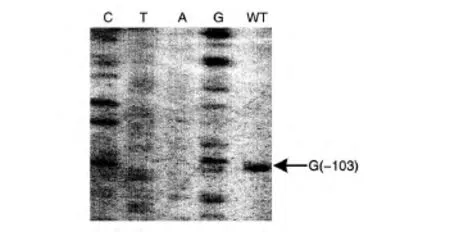
圖3 VPA1027的引物延伸實(shí)驗(yàn)結(jié)果

圖4 靶基因啟動子區(qū)序列
引物延伸和5′-RACE實(shí)驗(yàn)是尋找基因轉(zhuǎn)錄起始位點(diǎn)的兩個(gè)常用方法,二者各有優(yōu)缺點(diǎn)。引物延伸實(shí)驗(yàn)是一個(gè)逆轉(zhuǎn)錄的過程,其所得cDNA的量和mRNA的起始量成正比,因此它不僅可用來定位基因的轉(zhuǎn)錄起始位點(diǎn),還可用于研究調(diào)控子對靶基因的調(diào)控關(guān)系[1-5,14-16]。然而,在轉(zhuǎn)錄起始位點(diǎn)搜尋的過程中,需要針對每個(gè)待研究的基因設(shè)計(jì)多條引物進(jìn)行預(yù)實(shí)驗(yàn),費(fèi)時(shí)費(fèi)力費(fèi)財(cái)、成功率低,且易產(chǎn)生放射性污染,對實(shí)驗(yàn)者的健康也有損傷。5′-RACE實(shí)驗(yàn)雖然成功率高、避免了放射性同位素帶來的不便,但是它所得到的轉(zhuǎn)錄起始位點(diǎn)受mRNA降解片段的影響,易出現(xiàn)假陽性,而且也不能用于研究調(diào)控子對靶基因的調(diào)控關(guān)系。基于此,最合理的方法是將5′-RACE實(shí)驗(yàn)和引物延伸實(shí)驗(yàn)緊密結(jié)合起來,首先利用5′-RACE實(shí)驗(yàn)找出靶基因的轉(zhuǎn)錄起始位點(diǎn),再針對該位點(diǎn)設(shè)計(jì)特異性引物,利用引物延伸實(shí)驗(yàn)進(jìn)行驗(yàn)證,從而可以大大提高引物延伸實(shí)驗(yàn)的成功率。
[1]Zhang Y,Wang L,Han Y,et al.Autoregulation of PhoP/PhoQ and positive regulation of the cyclic AMP receptor pro?tein-cyclic AMP complex by PhoP in Yersinia pestis[J].J Bacteriol 2013,195(5):1022-1030.
[2]Zhang Y,Wang L,Fang N,et al.Reciprocal regulation of pH 6 antigen gene loci by PhoP and RovA in Yersinia pes?tis biovar Microtus[J].Future Microbiol,2013,8(2):271-280.
[3]Zhang Y,Qiu Y,Tan Y,et al.Transcriptional regulation of opaR,qrr2-4 and aphA by the master quorum-sensing regula?tor OpaR in Vibrio parahaemolyticus[J].PLoS One,2012,7(4):e34622.
[4]Sun F,Gao H,Zhang Y,et al.Fur is a repressor of biofilm formation in Yersinia pestis[J].PLoS One,2012,7(12):e52392.
[5]Zhang Y,Gao H,Wang L,et al.Molecular characterization of transcriptional regulation of rovA by PhoP and RovA in Yersinia pestis[J].PLoS One,2011,6(9):e25484.
[6]Yeung P S,Boor K J.Epidemiology,pathogenesis,and preven?tion of foodborne Vibrio parahaemolyticus infections[J].Food?borne Pathog Dis,2004,1(2):74-88.
[7]Lin Z,Kumagai K,Baba K,et al.Vibrio parahaemolyticus has a homolog of the Vibrio cholerae toxRS operon that medi?ates environmentally induced regulation of the thermostable di?rect hemolysin gene[J].J Bacteriol,1993,175(12):3844-3855.
[8]Whitaker W B,Parent M A,Boyd A,et al.The Vibrio para?haemolyticus ToxRS regulator is required for stress tolerance and colonization in a novel orogastric streptomycin-induced adult murine model[J].Infect Immun,2012,80(5):1834-1845.
[9]Boles B R,McCarter L L.Vibrio parahaemolyticus scrABC,a novel operon affecting swarming and capsular polysaccharide regulation[J].J Bacteriol,2002,184(21):5946-5954.
[10]Kim Y K,McCarter L L.ScrG,a GGDEF-EAL protein,par?ticipates in regulating swarming and sticking in Vibrio para?haemolyticus[J].J Bacteriol,2007,189(11):4094-4107.
[11]Trimble M J,McCarter L L.Bis-(3'-5')-cyclic dimeric GMP-linked quorum sensing controls swarming in Vibrio parahaemo?lyticus[J].Proc NatlAcad SciUSA,2011,108(44):18079-18084.
[12]Yu Y,Yang H,Li J,et al.Putative type VI secretion sys?tems of Vibrio parahaemolyticus contribute to adhesion to cul?tured cellmonolayers[J].Arch Microbiol,2012,194(10):827-835.
[13]張義全,高鶴,王麗,等.鼠疫菌H-NS蛋白的表達(dá)與純化及其DNA結(jié)合活性分析[J].微生物學(xué)報(bào),2011,51(5):615-621.
[14]Sun F,Zhang Y,Wang L,et al.Molecular characterization of direct target genes and cis-acting consensus recognized by quorum-sensing regulator AphA in Vibrio parahaemolyticus[J].PLoS One,2012,7(9):e44210.
[15]Ma L,Zhang Y,Yan X,et al.Expression of the Type VI se?cretion system 1 component Hcp1 is indirectly repressed by OpaR in Vibrio parahaemolyticus[J].Scientific World J,2012,2012:982140.
[16]Qu S,Zhang Y,Liu L,et al.Cyclic AMP receptor protein is a repressor of adenylyl cyclase gene cyaA in Yersinia pestis[J].Can J Microbiol,2013,59(5):304-310.
猜你喜歡
作文·小學(xué)低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學(xué)生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學(xué)低年級(2024年2期)2024-04-29 00:00:00
作文·小學(xué)低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(bào)(2022年4期)2022-08-09 08:52:06
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
