烏金豬FTO基因的克隆和表達分析
2014-10-27 09:04:38黃英楊明華畢保良潘洪彬俞政全高士爭李永能趙素梅
生物技術通訊 2014年3期
關鍵詞:分析
黃英,楊明華,畢保良,潘洪彬,俞政全,高士爭,李永能,趙素梅
云南農業大學 a.云南省動物營養與飼料重點實驗室;b.科技管理處;c.招生就業處;云南 昆明 650201
隨著經濟的快速發展和生活水平的不斷提高,肥胖已成為現代社會最常見的營養障礙性疾病之一,肥胖患病率呈現明顯上升的趨勢。肥胖可導致心血管疾病、高血壓、2型糖尿病和某些癌癥,從而增加死亡風險[1]。隨著對肥胖發生在基因水平上的深入研究,與肥胖相關的肥胖易感基因——脂肪和肥胖相關基因(fat mass and obesity associated gene,FTO基因)受到人們的關注。豬具有與人類相似的器官構成和代謝類型,尤其是在脂肪代謝和沉積方面,是研究人類肥胖的最佳模型。然而,豬FTO基因的相關研究十分欠缺,對烏金豬FTO基因的研究未見報道。烏金豬是云南省地方豬種之一,具有耐粗飼、適應性強、腹部脂肪沉積較多等性狀特點,是研究人類肥胖疾病機理的理想動物模型。
我們采用反轉錄PCR技術,從烏金豬脂肪組織中克隆FTO基因,并對該基因的序列結構和特征進行了生物信息學分析,同時通過軟件對該蛋白質的空間結構進行了預測,構建其分子系統進化樹,并采用實時PCR方法對7種組織進行了組織表達譜分析,為深入研究烏金豬FTO基因的生物學功能奠定了重要基礎。
1 材料與方法
1.1 材料
屠宰成年烏金豬3頭,取所需各種組織樣本,液氮速凍,-80℃保存。TRIzol-A+試劑購于北京天根生化科技有限公司;EasyScript第一鏈cDNA合成試劑盒購于北京全式金生物技術有限公司;pMD18-T載體、DL2000 DNA marker等均購于大連寶生物工程有限公司;其他試劑為國產分析純產品。
1.2 RNA提取
用TRIzol-A+試劑提取1頭烏金豬脂肪組織總RNA,用核酸蛋白定量儀測定RNA的濃度及純度,D260nm/D280nm=1.8~2.0,并經10 g/L瓊脂糖凝膠電泳鑒定其完整性后備用。
1.3 PCR引物設計及PCR反應條件優化
根據GenBank中公布的豬的相應基因序列(登錄號:NM_001112692.1),用Primer Primer 5.0軟件針對該基因設計2對引物,一對用于FTO基因的克隆,另一對用于FTO基因多組織的實時PCR,并以18S rRNA看家基因為內參。引物序列見表1,由生工生物工程(上海)有限公司合成。
FTO基因克隆PCR反應條件:95℃預變性5 min;95℃變性 40 s,58℃退火 40 s,72℃延伸 30 s,35個循環;72℃延伸10 min。用ddH2O代替反轉錄產物作為空白對照,用以檢驗是否有外源基因和基因組DNA污染。
1.4 cDNA合成與基因克隆
從1頭烏金豬脂肪組織中提取總RNA,根據所測定的濃度,各取2 μg總RNA進行反轉錄,用Ea?syScript第一鏈cDNA合成試劑盒進行反轉錄(42℃反轉錄30 min,85℃加熱5 min以使反轉錄酶失活),反應體系為20 μL。反轉錄后的cDNA單鏈可于-20℃保存或立即用于PCR。用生工生物工程(上海)有限公司的膠回收試劑盒回收PCR擴增的目的片段,并將其連接到pMD18-T載體上,轉入大腸桿菌,涂布LB培養基平板,長出菌落后經藍白斑篩選挑取白色單菌落,于LB培養液中振蕩培養12 h左右,分別取PCR鑒定過的菌液提取重組質粒,用KpnⅠ/HindⅢ雙酶切鑒定,將鑒定過的菌液及質粒送生工生物工程(上海)有限公司進行DNA序列測定。
1.5 生物信息學分析
將反轉錄PCR所得基因片段測序后,用DNA?Star 6.0軟件包中的Seqman程序進行序列拼接,用BLAST工具和DNAStar 6.0軟件包進行序列同源性比對和cDNA序列分析,用Meg4軟件進行系統進化樹分析;利用 ExPASy、SignalP、ProtScale、NetPhos、NetNGlyc、TMHMM、SMART、PSIPRED 和 SWISSMODEL網站在線程序對FTO蛋白的等電點、相對分子質量、信號肽、疏水性輪廓、磷酸化位點、糖基化位點、跨膜信息、二級結構及三級結構進行預測。所用相關分析軟件的生物信息學網址見表2。
1.6 實時PCR分析組織表達譜
分別提取3頭烏金豬心、肝、脾、肺、腎、肌肉和脂肪等7種組織的RNA,反轉錄為cDNA,以FTO F2/R2為引物進行實時PCR。反應條件:95℃預變性30 s;95℃變性5 s,60℃退火30 s,72℃延伸30 s,40個循環;72℃延伸10 min。18S rRNA和FTO基因熒光定量PCR反應條件只有退火溫度不同為55℃。每樣品3頭豬做3次重復,每個重復做2個平行,同時用ddH2O代替反轉錄產物和熒光試劑作為空白對照,用以檢驗是否有外源基因和基因組DNA污染。將純化的目的基因或內參基因的PCR產物進行1/10梯度稀釋后做標準曲線,共8個梯度濃度,每個濃度2個平行重復。相對表達量的分析以18S rRNA基因作為內參基因,通過參照基因表達水平校正待測樣本中目標基因的相對表達量,然后將各樣品的Ct值與標準曲線進行比較,得出其起始模板拷貝數。

表1 PCR引物

表2 生物信息學軟件及網址
2 結果
2.1 烏金豬脂肪組織總RNA的提取
取3 μL提取的總RNA經10 g/L瓊脂糖凝膠電泳分離,結果見圖1,28S和18S條帶清晰,且28S和18S灰度比約為2∶1,無脫尾現象出現,點樣孔無殘留,表明RNA無降解,質量可靠,可用于后續實驗。
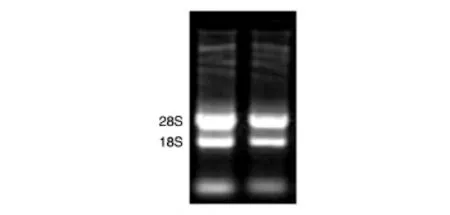
圖1 烏金豬脂肪組織總RNA的瓊脂糖凝膠電泳
2.2 FTO基因的克隆
反轉錄PCR得到1518 bp的片段,經10 g/L瓊脂糖凝膠電泳檢測,確認擴增產物與目的片段大小一致。將克隆的片段與pMD18-T載體連接,挑取陽性克隆,提取重組質粒,用KpnⅠ/HindⅢ雙酶切鑒定,表明目的基因FTO已成功插入載體。見圖2。
2.3 烏金豬FTO基因序列及對應蛋白
通過對PCR產物原液雙向測序及克隆后提取的陽性質粒雙向測序,確定了1518 bp的編碼區(CDS)序列(已提交GenBank,登錄號為JQ031263),編碼由505個氨基酸殘基組成的蛋白(圖3)。

圖2 PCR擴增FTO基因(A)及重組質粒的雙酶切鑒定(B)

圖3 烏金豬FTO基因編碼區序列及對應的氨基酸序列
2.4 生物信息學分析
2.4.1 同源性分析 用DNAMAN軟件將由FTO CDS推導的氨基酸序列與NCBI網站BLAST后下載的牛(登錄號:BC140478.1)、人(登錄號:NM_001080432.2)、羊(登錄號:EU072419.1)和大鼠(登錄號:BC168239.1)等4個物種的FTO CDS編碼的氨基酸序列進行比對,同源性分別為91%、89%、90%和83%(圖4)。
2.4.2 等電點與相對分子質量預測 利用ExPASy網站在線工具預測的FTO蛋白的理論等電點為5.18,相對分子質量為58.16×103。
2.4.3 進化樹分析 用Meg4軟件構建烏金豬、牛、人、羊和大鼠FTO蛋白的系統進化樹,結果顯示烏金豬與牛、羊的親緣性較近,其次是人、大鼠(圖5)。
2.4.4 FTO蛋白的特征分析 用TMHMM程序預測FTO蛋白的跨膜結構,發現該蛋白無跨膜區,表明該蛋白不是跨膜蛋白;用SignalP 3.0 Server軟件分析,發現該蛋白無信號肽出現;用NetPhos 2.0 Server對蛋白序列的磷酸化位點進行預測,發現烏金豬FTO蛋白結構中的磷酸化修飾位點共22個,包括10個絲氨酸蛋白激酶磷酸化位點、7個蘇氨酸蛋白激酶磷酸化位點、5個酪氨酸蛋白激酶磷酸化位點(圖6);用NetNGlyc 1.0 Server對FTO蛋白的糖基化位點進行預測,發現在200、246、302位氨基酸殘基處有糖基化位點(圖7);用網上在線工具ExPASy-ProtScale預測FTO蛋白的疏水性,發現該蛋白的疏水性最大值為3.056,最小值為-3.222,表明該蛋白是一個親水性蛋白(圖8)。

圖4 人(Homo)、大鼠(Rattus)、牛(Bos)、羊(Ovis)和烏金豬FTO CDS編碼的氨基酸序列的同源性比較
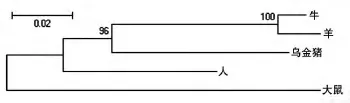
圖5 人、大鼠、牛、羊和烏金豬的FTO進化樹
2.4.5 FTO蛋白的二級結構和三級結構 用PSIPRED程序預測該蛋白的二級結構,結果表明共有201個螺旋(helix)、33個伸展鏈(strand)和271個卷曲結構(coil)(圖9)。用網上在線工具SWISSMODEL分析了該蛋白的三級結構(圖10)。
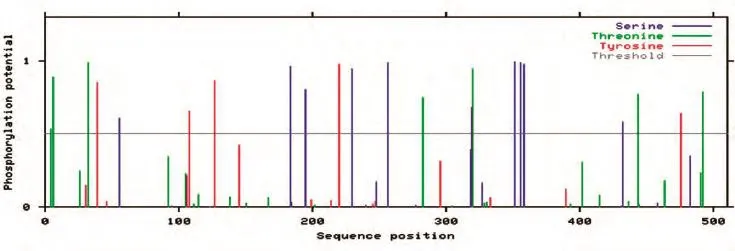
圖6 FTO蛋白的磷酸化位點預測
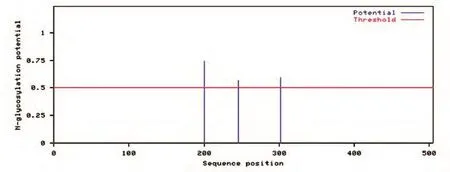
圖7 FTO基因的糖基化位點預測
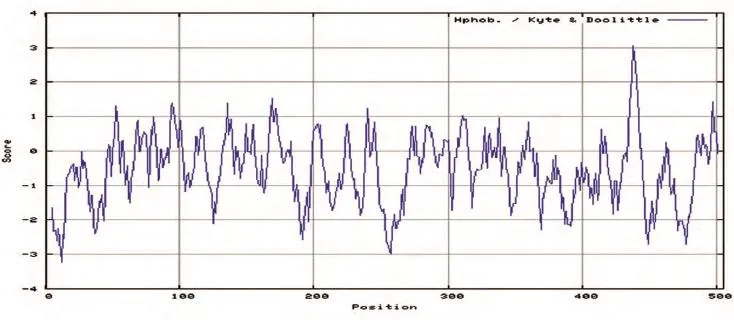
圖8 FTO蛋白的疏水性預測
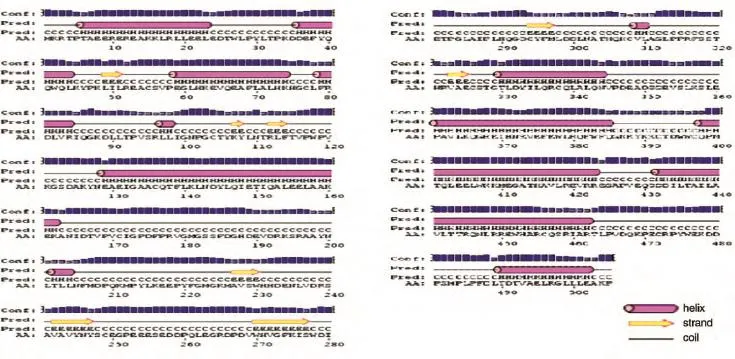
圖9 FTO蛋白的二級結構預測
2.5 FTO基因的組織表達譜
用FTO F2/R2引物分別擴增了3頭烏金豬的7種組織的cDNA,實時PCR結果表明,FTO基因mRNA在肝臟組織中的表達最高,在腎、脂肪、肺和脾組織中也大量表達,在心臟和肌肉組織中少量表達(圖11)。
3 討論
1999年,一篇關于腳趾融合小鼠的文獻對FTO基因進行了報道[2],但隨后的研究進展緩慢,直至2007年發現人的該基因與肥胖有關,才逐漸成為研究熱點。FTO基因是一個與肥胖相關的等位基因[3],屬于非血紅色加雙氧酶基因超家族,編碼2-酮戊二酸依賴的核酸脫甲基酶[4-5]。FTO廣泛存在于脊椎動物和海洋藻類中,是一個攜帶核定位信號的球狀蛋白。FTO基因mRNA在人、鼠大多數組織中均有表達,尤其在下丘腦中表達豐富[4,6]。本研究表明,FTO基因mRNA在烏金豬7個組織中的表達量有差異,在肝組織中表達量最高,脂肪、腎、脾中也有大量表達,心臟、肌肉中表達量最少。Frayling等用實時PCR發現FTO基因編碼蛋白在人體組織中廣泛分布,包括脂肪組織、胰島β細胞、垂體、腎上腺、大腦,尤其是在下丘腦弓狀核中高表達[3-4]。郭兵等研究表明,梅山豬FTO基因在海馬、脂肪、肝臟和肌肉組織中均有表達,在海馬中的表達量最高,在脂肪和肝臟組織中表達量較高,肌肉組織中的表達量較少[7]。
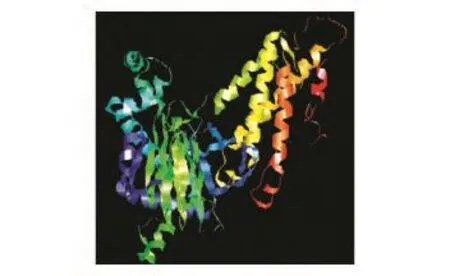
圖10 FTO蛋白的三級結構預測
我們克隆了烏金豬FTO基因編碼區全長系列,并和人、牛、羊和鼠等4個物種進行了進化樹分析,表明烏金豬FTO與牛、羊的親緣性較近,其次是人、大鼠。
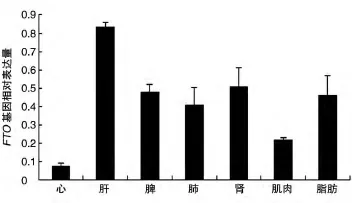
圖11 烏金豬FTO基因的組織表達譜
生物信息學是通過數學、統計學和計算機科學等學科對生物數據進行分析的方法和技術,由于它快速、高效和低成本,已被廣泛用于預測蛋白質的結構、功能和其他生物學特征[8]。蛋白質分析一般包括信號肽、跨膜結構和磷酸化位點、二級結構等的分析。蛋白質磷酸化是最常見、最重要的一種蛋白質翻譯后修飾方式[9],它參與和調控生物體內的許多生命活動,如信號轉導、基因表達、細胞周期調控等諸多細胞過程[10]。蛋白質磷酸化位點主要發生在絲氨酸、蘇氨酸和酪氨酸殘基上[11]。本研究表明烏金豬FTO蛋白結構中的磷酸化修飾位點共22個,這些磷酸化位點將是調控FTO蛋白功能的關鍵位點。糖基化是真核生物蛋白質翻譯后重要的修飾之一,有調節蛋白質功能的作用[12],我們推測的烏金豬FTO氨基酸序列在200、246、302位有糖基化位點。從氨基酸序列到蛋白質二級結構,是理解蛋白質結構和功能的重要一步,對蛋白質二級結構的預測有助于了解其三維構象和分析其結構域[13]。本研究結果表明,推測的烏金豬FTO氨基酸序列共有201個螺旋、33個伸展鏈和271個卷曲結構。蛋白質的結構與功能關系密切,以上關于FTO推測的氨基酸序列分析將為其蛋白質的生物學功能研究提供理論依據。
[1]Kopelman P.Health risks associated with overweight and obe?sity[J].Obesity Rev,2007,8(1):13-17.
[2]Peters T,Ausmeier K,Ruther U.Cloning of Fatso(Fto),a nov?elgene deleted by the Fused toes(Ft)mouse mutation[J].Mamm Genome,1999,10(10):983-986.
[3]Frayling T M,Timpson N J,Weedon M N,et al.A common variant in the FTO gene is associated with body mass index and predisposestochildhood and adultobesity[J].Science,2007,316(5826):889-894.
[4]Gerken T,Girard C A,Tung Y C L,et al.The obesity-asso?ciated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase[J].Science,2007,318(5855):1469-1472.
[5]Fernandez A M,Kim J K,Yakar S,et al.Functional inactiva?tion of the IGF-1 and insulin receptors in skeletal muscle causes type 2 diabetes[J].Genes Dev,2001,15:1926-1934.
[6]Fredriksson R,Hagglund M,Olszewski P K,et al.The obesi?ty gene,FTO,is of ancient origin,up-regulated during food deprivation and expressed in neurons of feeding-related nu?clei of the brain[J].Endocrine,2008,149(5):2062-2071.
[7]郭兵.豬FTO的組織特異性表達及其調控[D].南京:南京農業大學,2011.
[8]Romano P,Giugno R,Pulvirenti A.Tools and collaborative environmentsforbioinformatics research[J].BriefBioinform,2011,12(6):549-561.
[9]Kim J H,Lee J,Oh B,et al.Prediction of phosphorylation sites using SVMs[J].Bioinformatics,2004,20(17):3179-3184.
[10]姜錚,王芳,何湘,等.蛋白質磷酸化修飾的研究進展[J].生物技術通訊,2009,20(2):233-237.
[11]Blom N,Gammeltoft S,Brunak S.Sequence and structurebased prediction of eukaryotic protein phosphorylation sites[J].J Mol Biol,1999,294(5):1351-1362.
[12]Gupta R,Brunak S.Prediction of glycosylation across the hu?man proteome and the correlation to protein function[J].Pac Symp Biocomput,2002,7:310-322.
[13]張成崗,賀福初.生物信息學方法與實踐[M].北京:科學出版社,2002:158-230.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
