紅地球與夏黑無核成熟期光合日變化規律研究
2014-09-10 06:28:16聶松青石雪暉田淑芬劉昆玉楊國順鐘曉紅倪建軍
河北林業科技 2014年1期
聶松青 ,石雪暉 ,田淑芬 ,劉昆玉 ,楊國順 ,鐘曉紅 ,倪建軍 ,徐 豐 ,白 描
(1.湖南農業大學園藝園林學院,湖南 長沙 410128;2.湖南省葡萄工程技術研究中心,湖南 長沙 410128;3.天津市農科院葡萄研究中心,天津 300112)
葡萄葉片的光合特性和光合能力直接影響到葡萄漿果的品質和產量的形成,光合作用生成的有機化合物不僅為生物提供能量,也是生物用以建造自身軀體的原料。隨著便攜式光合測定系統的日益普及和對光合效率與作物產量關系認識的深入,人們越來越廣泛地把光合特性和光合效率用作選育和鑒定優良資源的重要指標。目前,紅地球葡萄是國內栽培面積最大的葡萄品種之一,夏黑無核葡萄因其早熟、粒大、無核等優良品性得以迅速推廣。在南方高溫高濕、避雨栽培環境下,研究紅地球和夏黑無核葡萄的光合特性,有利于采取合理的栽培措施和管理模式,對提高設施葡萄葉片光能的利用效率,提高果實的產量和品質具有重要意義[1]。
1 材料與方法
1.1 試驗材料
供試葡萄品種為湖南農業大學葡萄教學科研基地7a生紅地球和夏黑無核葡萄,南北行向,避雨栽培,架式為小平棚。選擇生長發育良好、生長勢基本一致的紅地球葡萄和夏黑無核葡萄品種各3株,每株選擇同側節位無病蟲害、長勢中庸的結果枝上第6節位功能葉作為測定葉片,每片葉重復測定3次。
1.2 試驗方法
在夏黑無核成熟期(7月上旬)和紅地球成熟期(8月下旬)的大晴天,使用美國LI-COR公司生產LI-6400XT便攜式光合儀,采用自然光源,于8:00~18:00每2h測定1次,每次重復3次。記錄的主要參數有凈光合速率(Pn,μmol·m-2·s-1)、光照有效輻射(PARi,μmol·m-2·s-1)、蒸騰速率(Ts,mmol H2Om-2·s-1)、氣孔導度(Gs,cm·s-1)、胞間CO2濃度(Ci,μmol·mol-1)、相對濕度(RH,%)、葉片溫度(Tleaf,℃)。
1.3 數據處理
采用Excel 2007處理數據及繪制圖表。
2 結果與分析
2.1 兩個葡萄品種成熟期環境因子的日變化
2.1.1 光合有效輻射(PAR)日變化
光合有效輻射對葡萄的生長發育影響很大,在一定光照強度范圍內,隨著光照強度的增加,光合效率也相應增加。在測定期間,從試驗數據發現(圖1),太陽光照強度日變化表現為先增加后減少的“單峰型”曲線,即清晨光強較弱,在上午 8:00~10:00增長迅速,午間 13:00左右最強,之后又逐漸減弱,回到上午8:00的光合有效輻射水平。
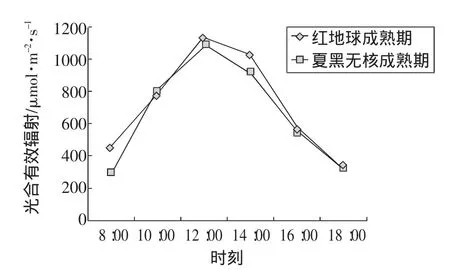
圖1 光合有效輻射日變化
2.1.2 葉片溫度(Tleaf)日變化
南方夏季炎熱,白天高溫天氣持續時間長。紅地球和夏黑無核葡萄的葉片氣溫日變化表現為先增加后減少的“單峰型”曲線變化趨勢,即早上8:00開始就有32℃左右的高溫,隨著時間的推移,溫度增加迅速,午間14:00左右最高,達到最高溫41.9℃(紅地球)和41.4℃(夏黑無核),午后又逐漸降低(圖2)。下午18:00,光強逐漸減弱,但平棚內溫度并未大幅降低,保持在37℃左右。
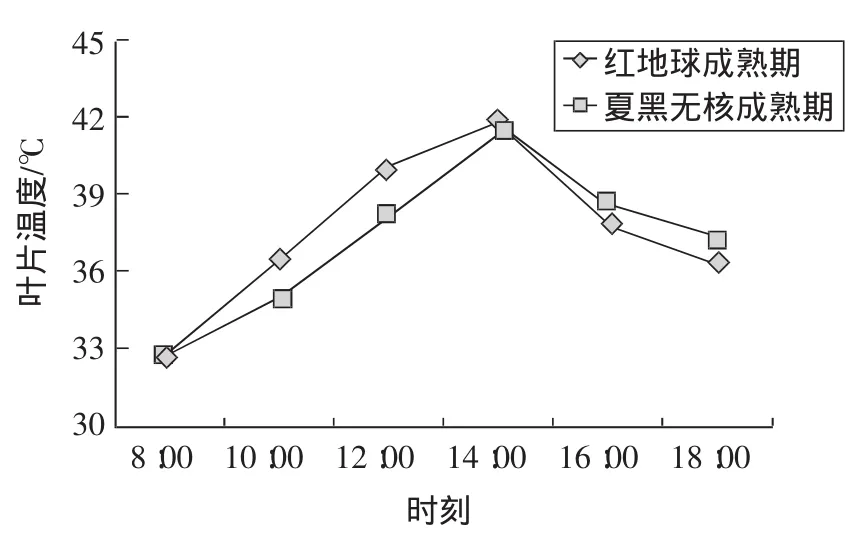
圖2 葉片溫度日變化
2.1.3 相對濕度(RH)日變化
葉片的濕度受氣溫和光照影響明顯。如圖3所示,紅地球和夏黑無核葡萄葉片相對濕度日變化表現為先下降后上升的趨勢。一天當中,設施內空氣相對濕度變化范圍為30%~50%。上午8:00的空氣濕度較大,隨著時間的遞增,空氣濕度逐漸變小,中午14:00左右空氣濕度最小,14:00過后空氣濕度有所增加。主要原因是上午溫度相對較低,水分蒸發速度較慢;中午溫度高,水分蒸發快;午后光合作用和蒸騰作用又開始增強,空氣濕度又有所增加,而后隨著溫度的降低和光照的減弱逐漸減小。空氣相對濕度的變化趨勢與溫度變化趨勢相反,可能與大氣溫度、光強等環境因子直接相關。
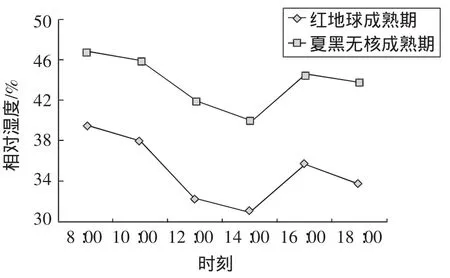
圖3 相對濕度日變化
2.2 兩個葡萄品種成熟期光合生理因子的日變化
2.2.1 胞間CO2濃度(Ci)日變化
隨著時間的推移,紅地球葡萄胞間CO2濃度呈現出“W型”曲線變化,夏黑無核葡萄呈現“V型”曲線變化。清晨,葉片氣孔敞開,大量吸收大氣CO2而凈光合速率較低,固定CO2能力較低,使細胞間隙CO2濃度最高;隨著溫度、有效光輻射的升高,較多的CO2被固定,引起細胞間隙CO2濃度下降。隨著氣溫升高和光照增強,10:00左右當凈光合速率最高時,紅地球葡萄胞間CO2濃度下降到第1個低值,夏黑無核細胞間隙CO2濃度達到最低值。光合“午休”時紅地球葡萄因氣孔關閉阻止細胞內外CO2濃度交換,胞間CO2濃度相對較低,而此時夏黑無核葡萄濃度沒有降低反而上升是因為有非氣孔限制因素。午后,雖然葉片氣孔再次敞開,紅地球葡萄胞間CO2濃度迅速回升。此后隨著時間推移,光合作用減弱,胞間CO2濃度又有所增加,如圖4。
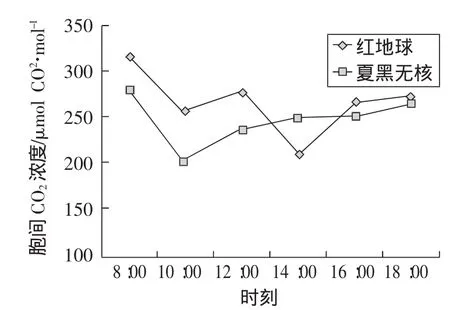
圖4 胞間CO2濃度日變化
2.2.2 氣孔導度(Gs)日變化
在光照強度較低的情況下,氣孔導度(Gs)隨光照強度的升高而增大,隨CO2濃度的升高而減小。在不同光照強度下,氣孔導度隨CO2濃度變化的幅度有所不同,但總的來說都是隨著二氧化碳濃度的增加而減小。由圖5可知,紅地球和夏黑無核葡萄葉片氣孔導度日變化規律是雙峰曲線。從早上8:00開始,隨光強的增加和氣溫的上升,氣孔逐漸打開,蒸騰作用加強,第一個峰值都出現在上午10:00左右,隨著時間的推移逐漸下降。午間的高溫強光使葉溫上升,氣孔阻力增大,氣孔導度迅速減小,在下午14:00左右出現低谷,之后氣溫回落,在一定適宜溫度范圍內,氣孔再度張開,氣孔導度又經過短期的增長到16:00左右出現第2個峰值,之后隨著溫度的下降氣孔導度逐漸減小呈下降趨勢。
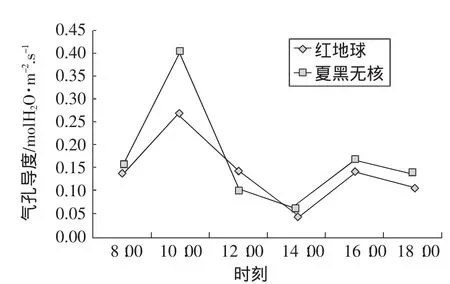
圖5 氣孔導度的日變化
2.2.3 蒸騰速率(Tr)日變化
蒸騰速率取決于葉片內外的蒸氣壓差和擴散途徑阻力的大小,光照、溫度、大氣相對濕度是影響蒸騰速率的主要因素。氣孔導度和蒸騰速率的變化趨勢基本一致,說明蒸騰速率在很大程度上決定于氣孔的活動狀態。上午8:00左右,葉片氣孔導度較小,蒸騰作用較弱。隨著光照增強,氣溫逐漸增高,葉片水汽壓差也隨之增大,蒸騰速率不斷升高,10:00左右出現第1次峰值;之后隨著時間推移,光強增大,氣溫升高,葉片氣孔逐漸縮小直至關閉,蒸騰速率隨之降低;午后“午休”過后,蒸騰作用隨光合作用增強而增強,16:00左右達到第2次峰值,此后蒸騰速率隨時間推移遞減(圖 6)。
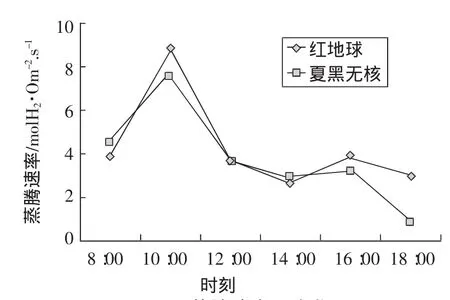
圖6 蒸騰速率日變化
2.2.4 凈光合速率(Pn)日變化
晴朗天氣條件下,紅地球和夏黑無核葡萄葉片成熟期Pn日變化趨勢雙峰曲線。在光飽和點以下,隨時間的推移和光合有效輻射的增強,光合速率急劇增大,第1次峰值出現在上午10:00左右,達到一天中最大凈光合速率(即紅地球Pnmax 為 12.8μmol/m2·s和夏黑無核 Pnmax為11.8μmol/m2·s)。而后由于光照過強,引起氣溫升高,空氣濕度下降,導致葉片水熱不協調,進而導致氣孔收縮,氣孔阻力增大,氣孔導度迅速減小,CO2吸收量減小,光合速率下降。在高溫、低CO2濃度的午間,產生光抑制現象,致使光合作用減弱,14:00左右葉片出現明顯的光合“午休”現象。午后隨著氣溫降低,在一定溫度范圍內,光合速率又緩慢回升,第2次峰值出現在下午16:00左右。第2峰值相對第1峰值較低。之后,隨著溫度的下降光合速率逐漸減小。
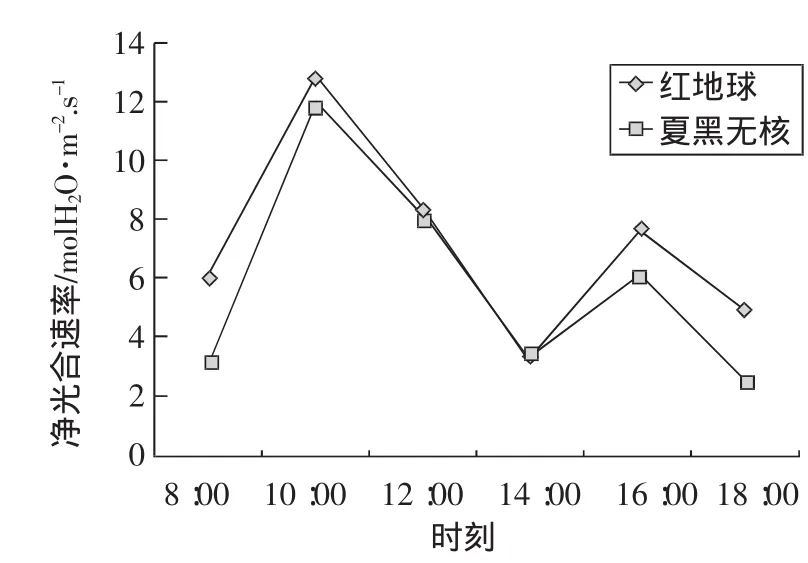
圖7 紅地球與夏黑無核葡萄凈光合速率日變化
2.3 關于光合日變化的影響因子及相關性
本研究發現,在果實成熟期,無論是紅地球,還是夏黑無核葡萄,環境因子光合有效輻射和葉片溫度均呈先升后降的“單峰”型曲線變化趨勢,而相對濕度呈相反的變化趨勢,即先下降后上升再下降。兩種葡萄在成熟期溫度變化受光照強度的影響,兩者變化呈正相關;而濕度與光照變化趨勢相反,呈負相關。光照強度是影響溫度和濕度的主要因子,同時三者的相互作用又對葉片的光合特性產生影響。
兩個葡萄品種葉片凈光合速率(Pn)與主要生理生態因子之間存在密切關系,凈光合速率(Pn)分別與葉片蒸騰速率(Tr)和氣孔導度(Gs)呈極顯著正相關。蒸騰速率、氣孔導度與光合速率日變化趨勢相似且升降幅度相對較大,且一天中的最大值為第1峰值。
3 討論與結論
植物光合作用的日變化類型主要有4種:正規曲線型、平坦型、變動型和中午降低型[2]。在果樹上主要有3種表現:單峰型曲線[3-4]、雙峰型曲線[5-6]和極少數的三峰型曲線[7]。強光、高溫、低濕、蒸騰速率高、氣孔關閉等生理生態因子可能是午間葉片出現光合“午休”現象的原因。Pn中午降低的因素主要有2個:氣孔因素和非氣孔因素。氣孔因素是由于中午光照、溫度和濕度等環境因子變化引起植物氣孔部分關閉,CO2進入葉片受阻而使光合下降,而非氣孔因素是由于葉肉細胞自身羧化酶活性的下降而引起Pn降低。根據Farquhar和Sharkey[8]的觀點,只有當 Pn和 Ci二者同時減小,且Ls(1-Ci/Ca)降低時,才可以認為Pn下降主要是由氣孔引起的,否則Pn下降要歸因于葉肉細胞羧化能力的降低。通過對成熟期紅地球葡萄的Ci、Ls與Pn之間的關系進行分析,Ls在進入光合“午休”階段呈下降的趨勢,此時Ci、Pn均呈下降的變化。表明紅地球葉片光合“午休”導致Pn下降的主要原因均是氣孔因素。同時通過對成熟期夏黑無核葡萄的Ci、Ls與Pn之間的關系進行分析,可推斷出,Ls在進入光合“午休”階段呈下降的趨勢,而此時Ci呈上升、Pn下降的變化,表明夏黑無核葉片光合“午休”導致Pn下降的主要原因均是非氣孔因素。
葡萄的生長發育與光合作用息息相關,葡萄光合作用合成的有機物不僅為漿果發育提供能量來源,也是其建造自身軀體的原料。光合生理生態因子之間都是相互聯系相互作用的,眾多因子間的相互作用對光合作用的影響至今都不清楚。今后,有關光合作用與環境因子的關系還需進一步探索研究。
[1]陳沖,楊國順,石雪暉,等.避雨栽培條件下紅寶石無核葡萄光合特性初探[J].中外葡萄與葡萄酒,2011,(3):21-24.
[2]呂忠恕.果樹生理[M].上海:上海科學技術出版社,1982:55.
[3]彭少兵,郭軍戰.不同樹莓和黑莓品種的光合特性研究[J].西北農林科技大學學報:自然科學版,2007,35(3):116-120.
[4]張志華.核桃光合特性的研究[J].園藝學報,1993,20(4):319-323.
[5]李延菊,李憲利.3個扁桃品種的光合特性[J].林業科學,2006,42(11):23-28.
[6]郭春芳,孫云,陳常頌,等.茶樹品種光合與水分利用特性比較及聚類分析[J].作物學報,2008,34(10):1797-1804.
[7]王文江.大磨盤柿樹光合特性的研究[J].園藝學報,1993,20(2):105-110.
[8]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Ann Rev PlantPhysiology,1982,33:317-345.
