神經生長因子對小鼠睪丸支持細胞生理狀態影響初探
2014-08-25 03:41:38付璐璐鄭連文徐耀宏張靜文梁文君高利偉胡文宇王春強
中國實驗診斷學 2014年3期
付璐璐,鄭連文,徐耀宏,張靜文,梁文君,高利偉,胡文宇,王春強
(1.吉林大學第二醫院 婦產科生殖中心,吉林 長春130041;2.赤峰市醫院,內蒙古 赤峰024000;3.吉林大學動物科學學院,吉林 長春130000;4.遼寧醫學院,遼寧 錦州210700)
神經生長因子(NGF)是神經系統最重要的生物活性分子之一,在神經細胞的生長發育、神經系統正常功能的發揮等方面起著重要作用[1]。除作用于神經系統外,對心血管系統、免疫系統、生殖系統等也有影響,很早就有研究表明哺乳動物的睪丸、附睪、前列腺中有NGF及其受體存在[2]。NGF對哺乳動物生殖系統的影響是近年來的研究熱點之一,本研究將NGF對小鼠支持細胞的作用進行初步探索,為進一步闡明NGF對雄性動物睪丸發育及精子發生過程的調控機制提供科學證據。
1 材料與方法
1.1材料一周齡小白鼠、膠原酶(0.5 mg/ml)、胰酶(0.5 mg/ml)、PBS(0.3 mg/ml)、混合酶消化液 (0.1% collagenase,0.4% hyaluronidase,0.08% Dnase I和0.03% trypsin inhibitor)、培養基、顯微鏡、濾網、鑷子、剪刀、蒸餾水等。
1.2方法(1)2個睪丸去膜后,放入10 ml PBS中洗兩次,沉底;(2)將曲細精管加入4 ml膠原酶(0.5 mg/ml)中, 80 轉/min,33 ℃,10-15 min,沉底;(3)放入10 ml PBS中洗兩次,加入4 ml胰酶(0.5 mg/ml)消化5-10 min,34℃不搖;(4)放入10 ml PBS中洗兩次,用含4 ml trypsin inhibitor的PBS(0.3 mg/ml)洗一次.沉底(2 min);(5)加入4 ml混合酶消化液(0.1% collagenase,0.4% hyaluronidase,0.08% Dnase I和0.03% trypsin inhibitor) 80 轉/min,34℃,40 min;(6)離心500 rpm,4 min,放入3 ml PBS中洗兩次;(7)低滲純化SCs:1 ml PBS懸浮,加2.5 ml的1∶10稀釋的PBS,共3.5 ml,顛倒混3次,離心500 rpm,4 min;(8)10 ml的DMEM/F12洗兩次,10 ml重懸,計數后鋪板;(9)支持細胞的形態學觀察及HE染色;(10)油紅O染色鑒定睪丸支持細胞;(11)置于37℃、5%CO2、飽和濕度下培養,并隨時觀察兩個培養皿中細胞的生長情況。
2 結果
2.1HE染色結果顯示支持細胞凸起較多,細胞核呈類圓形,核仁清晰,胞漿淡染,細胞質內可見大小不一的空泡。
2.2油紅O染色顯示HE染色后的空泡結構被油紅染為橘黃色,大小不等,呈懸浮狀態,其實質為脂質小滴,散落在支持細胞中(見圖1)。
2.3 48小時后對照組體外培養的小鼠睪丸支持細胞未見明顯生長,NGF組小鼠睪丸支持細胞3-4小時生長即可以貼壁,48小時候已經長滿培養皿(見圖2)。

圖1 油紅O鑒定支持細胞的純度(×400)
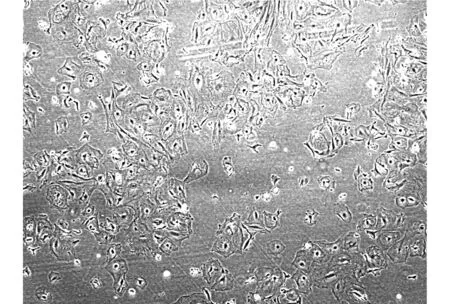
圖2 NGF組培養48小時后支持細胞(×100)
3 討論
近年來,人們在神經營養素對雄性動物繁殖系統作用的研究上取得了一定的進展。其中, NGF 的作用研究的最早,也較為系統。人們最先在小鼠的睪丸中發現了片段長度分別為 1.3 kb和1.5 kb的NGF的兩種轉錄產物。它們均存在于小鼠睪丸中的精母細胞和早期精原細胞中。如果把大鼠睪丸支持細胞、間質細胞和管周細胞分離培養數天后,仍可以在細胞中檢測出有大量NGF的表達。因此,可以得出結論,上述3種細胞均可以表達并分泌NGF[3]。在新生小鼠睪丸內,NGF的高親合力受體TrkA在生殖細胞中沒有表達,但表達于睪丸間質細胞中[4]。此后,在小鼠和人睪丸中的報道中也發現了類似的結果[5]。說明NGF可以通過自分泌或旁分泌的方式影響哺乳動物睪丸的發育,進而影響精子的發生。關于NGF在睪丸中的作用途徑,近年來,人們也進行了大量的研究。研究發現,生殖激素是調節NGF在哺乳動物睪丸功能發揮的關鍵。大鼠的垂體被摘除后,大鼠曲細精管中NGF受體TrkA 的表達量顯著降低,有些細胞甚至不再表達TrkA,說明垂體分泌的促性腺激素可以通過抑制TrkA表達影響NGF的功能的發揮[6]。此外,通過向大鼠體內注射hCG12h后,睪丸細胞內的TrkA水平顯著的提高。核糖核酸酶保護實驗表明,TrkA的mRNA水平在VⅡ和VⅢ期的表達最高[7];睪酮也能影響 TrkA 的表達。當睪丸間質細胞被破壞或雄激素受體被阻斷后,TrkA 的 mRNA 水平顯著的提高,然而,睪酮的處理卻會抑制 TrkA的表達[6]。以上說明雄激素可以調節睪丸中 NGF/TrkA 的信號傳遞。4-甲基兒茶酚(4-methylcatechol,4-MC)是促進NGF合成的一種兒茶酚類衍生物。當用10 μg/kg的4-MC 連續處理成年大鼠10天后,可顯著的提高血漿中NGF的水平,同時,能表達TrkA的組織中TrkA 的表達量都顯著提高。在成年的大鼠中,精母細胞、精細胞和精子中均有TrkA的表達,但是從表達量來看,TrkA 主要表達于精母細胞中。若用 4-MC處理新出生的大鼠,TrkA 的表達規律則與成年大鼠不同[8]。此外,人們通過建立TrkA敲除小鼠模型進一步說明了NGF/TrkA系統在雄性生殖生理中發揮的重要作用。TrkA被敲除掉后,胚胎期的第14天(E14)小鼠的睪丸中的曲細精管數目顯著的低于對照組。從組織學上也能看出睪丸的發育明顯的滯后。到了胚胎期的第19天(E19),TrkA敲除小鼠的睪丸曲細精管中的精原細胞的數目顯著減少,且曲細精管中的凋亡細胞的比例要比正常小鼠中多10倍以上[9]。然而,由于TrkA敲除小鼠出生后不久就會死亡,所以無法利用該模型分析NGF/TrkA系統對小鼠精子發生的影響。但是可以通過生精細胞的體外培養模型對此進行研究。當用NGF連續處理生精細胞5天后,生精細胞體外培養系統中處于第二次減數分裂后期的精母細胞的比例會顯著增加,而精細胞的數量顯著減少。但是,若用TrkA的抑制劑K252a處理后,精細胞的數量顯著增加[10]。在精子的發生過程中,間質細胞與支持細胞均在生殖細胞的成熟、分化和精子的生成中發揮重要作用[11]。因此,NGF 及其受體 TrkA 在睪丸中各種細胞的表達及分布,說明 NGF 有可能是通過旁分泌的方式影響著生精細胞的分化。當用NGF處理14天后,大鼠睪丸中的支持細胞特異的雄激素結合蛋白(androgen binding protein,ABP)可顯著的提高,說明在睪丸發育過程中ABP的作用可能受到NGF的調節[12]。在體外培養體系中,用10ng/mL 的NGF處理12天,可提高睪丸支持細胞的存活率,而用BDNF、NT-3和NT-4處理則沒有這種變化[13]。此外, NGF 還可以提高小鼠睪丸間質瘤細胞中類固醇激素的分泌[14]。本文實驗結果表明48小時后對照組體外培養的小鼠睪丸支持細胞末見明顯生長,NGF組小鼠睪丸支持細胞3-4小時生長即可以貼壁,48小時已經長滿培養皿。可知NGF可能促進小鼠睪丸支持細胞的生長。近年隨著人們對NGF的深入研究,發現其并不是神經系統所特有的分子蛋白,NGF及其受體廣泛存在于全身各個系統,并且發揮作用。
近年來的研究還發現,NGF 和 TrkA 同樣還存在于成熟的牛和人的精子中,并且在少弱精癥(oligoasthenozoospermic)患者的精液中 NGF 蛋白的含量和精子中 TrkA 的 mRNA水平均很低[15]。在牛成熟精子中,NGF 主要位于精子的頂體帽,核和尾部。若用 NGF處理2 h可以顯著的影響牛成熟精子瘦素及胰島素的分泌,并影響精子的活率和凋亡[16]。同時,NGF 還可以通過 MAPK 信號通路影響倉鼠精子的頂體反應,且這個過程依賴于 NGF的濃度和處理的時間[15]。
參考文獻:
[1]王占友,石玉秀.神經生長因子與運動神經再生[J].解剖科學進展,1997,3(2):126.
[2]Hater GP,Glanville RW,Thoenen H.The purif ication of nerve growthfactor from bovine seminal plasma[J].J Biol Chem,1982,275(14):8541.
[3]Jin W,Arai KY,Shimizu K,et al.Cellular localization of NGF and its receptorstrkA and p75LNGFR in male reproductive organs of the Japanese monkey,Macaca fuscata fuscata[J].Endocrine,2006,29(3):155.
[4]Djakiew D,Pflug B,Dionne C,et al.Postnatal expression of nerve growthfactor receptors in the rat testis[J].Biology of Reproduction,1994,51(2):214.
[5]Muller D,Davidoff MS,Bargheer O,et al.The expression of neurotrophins andtheir receptors in the prenatal and adult human testis:evidence for functions inLeydig cells.Histochemistry and Cell Biology,2006,126(8):199.
[6]Persson H,Ayer-Le Lievre C,So¨der O,et al.Expression of b-nerve growth89factor receptor mRNA in Sertoli cells downregulated by testosterone[J].Science,1990,247(7):704.
[7]Schultz R,Metsis M,Ho¨ kfelt T,et al.Expression of neurotrophin receptors inrat testis.Upregulation of TrkA mRNA with hCG treatment[J].Molecular andCellular Endocrinology,2001,182(6):121.
[8]Levanti MB,Germana` A,de Carlos F,et al.Effects of increased nerve growthfactor plasma levels on the expression of TrkA and p75 in rat testicles[J].Journalof Anatomy,2006,208(6):373.
[9]Cupp AS,Tessarollo L,Skinner MK.Testis developmental phenotypes inneurotropin receptor trkA and trkC null mutations:role in formation ofseminiferous cords and germ cell survival[J].Biology of Reproduction 2002,66(5):1838.
[10]Perrard M,Vigier M,Damestoy A,et al.b-Nerve growth factor participates inan auto/paracrine pathway of regulation of the meiotic differentiation of ratspermatocytes[J].Journal of Cellular Physiology,2007,210(3):51.
[11]Svechnikov K,Izzo G,Landreh L,et al.Endocrine disruptors and Leydig cellfunction[J].Journal of Biomedicine Biotechnology,2010,2010(1):1.
[12]Lonnerberg P,Soder O,Parvinen M,et al.b-Nerve growth factor influencesthe expression of androgen-binding protein messenger ribonucleic acid in therat testis[J].Biology of Reproduction,1992,47(8):381.
[13]Chen Y,Dicou E,Djakiew D.Characterization of nerve growth factorprecursor protein expression in rat round spermatids and the trophic effects ofnerve growth factor in the maintenance of Sertoli cell viability[J].Molecular andCellular Endocrinology,1997,127(2):129.
[14]Muller D,Davidoff MS,Bargheer O,et al.The expression of neurotrophins andtheir receptors in the prenatal and adult human testis:evidence for functions inLeydig cells[J].Histochemistry and Cell Biology,2006,126(8):199.
[15]Jin W,Tanaka A,Watanabe G,et al.Effect of NGF on the motility andacrosome reaction of golden hamster spermatozoa in vitro[J].Reproduction and Development,2010,56(3):437.
[16]Li C,Zheng L,Wang C,et al.Absence of nerve growth factor and comparisonof tyrosine kinase receptor A levels in mature spermatozoa fromoligoasthenozoospermic,asthenozoospermic and fertile men[J].Clinica ChimicaActa,2010,411(4):1482.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
