右美托咪定對感染性休克大鼠脾臟及胸腺caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的影響
2014-08-15 10:07:42秦培順黃蔥蔥
實用藥物與臨床 2014年12期
姬 斌,秦培順,南 洋,黃蔥蔥,李 軍
0 引言
感染性休克亦稱膿毒性休克,主要由革蘭陰性菌及該菌產生的內毒素過度刺激機體的免疫系統而產生過量炎性細胞因子[1]。感染性休克是臨床危重患者常見死亡原因,盡管有多種先進的治療方法,但在重癥病房的致死率依然高達20%以上[2]。目前研究成果表明,感染性全身炎癥反應患者的炎癥細胞因子和免疫蛋白主要在脾臟巨噬細胞內生成,胸腺組織中也有體現[3]。本次研究采用內毒素制備感染性休克大鼠模型,采用右美托咪定予以干預,觀察大鼠血清IL-4、IL-10、IFN-γ和IL-2的水平,并檢測脾臟及胸腺caspase-3、caspase-8和caspase-9活性及淋巴細胞凋亡情況。
1 資料與方法
1.1 實驗動物模型制作 選取雄性12周齡Walse大鼠36只(由溫州醫科大學實驗動物中心提供,許可證號:SYXY(浙)2009-0129),每籠3只。適應性喂養3 d后,采用數字表法隨機分為3組:空白組(S組)、內毒素組(E組)和右美托咪定治療組(D組)。S組:靜脈持續泵入0.9%生理鹽水1.0 mL/(kg·h);E組:靜脈注射內毒素15 mg/kg,2 min后持續泵入0.9%生理鹽水,劑量1.0 mL/(kg·h);D組:靜脈注射內毒素15 mg/kg,2 min后持續泵入右美托咪定(鹽酸右美托咪定注射液,商品名:艾貝寧,規格:200 μg/2 mL,江蘇恒瑞醫藥股份有限公司,批號:H20110086),劑量:5 μg/(kg·h)。
1.2 研究方法 用藥0.5、1、2、3、4、6 h后,每組選取存活Walse大鼠6只,頸動脈采血1 mL[2],檢測血清中IL-4、IL-10、IFN-γ和IL-2等炎性細胞因子水平,6 h后存活者以脫頸法處死,分離脾臟及胸腺組織,用10%福爾馬林溶液固定后,石蠟包埋、切片,于-70 ℃環境保存。
1.2.1 炎性細胞因子檢測 采用ELISA(酶聯吸附法)檢測3組大鼠血清IL-4、IL-10、IFN-γ和IL-2的含量,儀器選用μ-Quant酶標分光光度儀(美國Bio-Tek公司)、ELISA檢測試劑盒(BIoSoURCE公司)。
1.2.2 脾臟及胸腺caspase-3、caspase-8和caspase-9活性檢測 采用免疫組織化學染色法檢測。將脾臟、胸腺石蠟切片分別脫蠟、內源性過氧化物酶封閉后,分別滴加兔抗鼠caspase-3、caspase-8和caspase-9多克隆抗體(北京中杉金橋生物技術有限公司,滴度1∶100),PBS代替一抗做陰性對照。40 ℃過夜后滴加二抗,二抗為生物素標記羊抗小鼠1gG(Santa Cruz公司,滴度1∶200),DAB(美國Sigma公司)顯色,蘇木素復染,脫水,透明,封片[4]。采用彩色病理圖像分析系統對切片進行圖像分析。
1.2.3 淋巴細胞凋亡檢測 取脾臟、胸腺石蠟切片進行脫蠟、粉碎處理,經400目不銹鋼篩網過濾制成單細胞懸液,1 500 r/min離心5 min,棄上清,PBS漂洗2次,重懸PBS溶液中室溫孵育30 min后加入碘化丙啶(PI)染色,400目尼龍網再次過濾后,采用FACS Calibur流式細胞儀(美國Beckman公司)檢測,操作按說明書要求進行。
1.2.4 淋巴細胞亞群的檢測 取脾臟、胸腺單細胞懸液PBS漂洗2次,采用雙管標記檢測法,即采用CD4、CD8和IFN-γ FITC三色標記一管檢測Th1細胞,CD4、CD8和IL-4 PE三色標記一管檢測Th2細胞。室溫孵育15 min、SBP洗滌2次后,每管分為2份,分別加入APC標記的anti-mouse IFN-γ和IL-4與各自的同型對照,計算Th1(CD4+/IFN-γ+)和Th2(CD4+/IL-4′)的百分率。
1.2.5 圖像分析 采用Image-pro PLUS 6.0彩色病理圖像分析系統對切片進行圖像分析,采用陽性細胞計數法,每張切片在高倍鏡下隨機選取4個無重疊視野,計數每個視野的陽性細胞數,取平均值記錄。
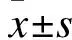
2 結果
2.1 三組大鼠不同時間點血清中各細胞因子濃度情況 與S組相比,E組、D組大鼠血清的抗炎因子IL-4、IL-10水平和促炎因子IFN-γ、IL-2均顯著提高,差異有統計學意義(P<0.05);隨治療時間延長,E組的IL-4、IL-10水平降低,IFN-γ、IL-2水平升高;D組的IL-4、IL-10水平升高,IFN-γ、IL-2水平降低,兩組間比較差異有統計學意義(P<0.05)。見表1。
2.2 三組大鼠脾臟caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡情況 與S組相比,E組、D組的大鼠脾臟caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數均顯著增多,差異均有統計學意義(P<0.05);與E組相比,D組大鼠脾臟caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數均顯著減少,差異有統計學意義(P<0.05),且caspase-9表達的陽性細胞數減少幅度最為明顯。見表2。
2.3 三組大鼠胸腺caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡情況 與S組相比,E組、D組的大鼠胸腺caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數均顯著增多,差異有統計學意義(P<0.05);與E組相比,D組大鼠胸腺caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數均顯著減少,差異有統計學意義(P<0.05),且caspase-9表達的陽性細胞數減少幅度最為明顯。見表3。

表1 三組大鼠不同時間點血清中各細胞因子濃度比較(ng/L)

表2 三組大鼠脾臟caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數比較(個/400倍野)

表3 三組大鼠脾臟caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數比較(個/400倍野)
2.4 E組和D組的Th1/Th2比率比較 D組大鼠脾臟及胸腺Th1/Th2比率均顯著高于E組,兩組的脾臟及胸腺Th1/Th2比率比較差異均有統計學意義(P<0.05)。見表4。

表4 E組和D組的Th1/Th2比較(%)
3 討論
IFN-γ和IL-2屬于促炎因子,IFN-γ主要由Thl與Tcl細胞分泌,IL-2由Thl細胞分泌;IL-4和IL-10屬于抗炎因子,主要由Th2細胞分泌[5]。發生感染性休克后,機體內的炎性反應體系迅速啟動,多種炎性細胞因子被釋放進入微循環[6]。現有研究結果表明,感染可以使免疫應答或向Thl或向Th2極化,這對成功控制感染和避免免疫能力下降的修復都非常重要[7]。Nikonenko等[8]認為,促炎因子介導細胞免疫反應,誘導 Thl細胞產生 IFN-α,增大 T淋巴細胞和 NK細胞的細胞毒作用。Boyman等[9]認為,IL-2水平與T淋巴細胞的免疫功能呈正相關,IL-2水平下降,T淋巴細胞的免疫清除能力就減弱,導致免疫損傷。Foitzik等[10]的研究則顯示,IL-10的抗炎作用在機體內十分普遍,貫穿炎癥反應的所有環節,其抑制炎癥、維持細胞因子平衡的作用已被眾多學者認可。而IL-4具有顯著的抗炎作用,可以有效抑制內毒素(LPS)介導的炎性因子產生[11],Th2細胞也主要通過IL-4參與機體免疫反應,調節機體免疫系統進行不同種類的免疫應答反應[12]。
炎性反應早期,促炎因子處于優勢,引發淋巴細胞凋亡,導致caspase-8聚合而發生激活,隨機產生caspase蛋白酶解級聯反應,并進一步激活其同源酶。caspase-3通常是以無活性前體存在于細胞中,當細胞進入凋亡時才被激活;caspase-8的級聯反應也能夠促使caspase-3激活[13]。細胞色素C(Cyt C)在細胞凋亡早期從線粒體膜間隙進入細胞漿中參與激活caspase-9,再激活caspase-3發生鏈式生物效應,最終使細胞DNA斷裂、細胞開始凋亡。激活后的caspase能再次促使線粒體釋放Cyt C,使細胞凋亡形式更加嚴重[14]。右美托咪定是一種高選擇性α腎上腺素受體激動藥[15],具有減輕炎癥介質的釋放和部分抑制炎癥因子的放大效應,使去甲腎上腺素的釋放減少,而去甲腎上腺素的水平具有調節機體免疫應答和炎癥反應的重要作用。因此,右美托咪定可能是通過減少去甲腎上腺素的釋放而使炎癥反應減輕,并改善免疫能力[16]。
本研究采用右美托咪定干預治療感染性休克大鼠,結果IL-4、IL-10水平隨治療時間延長而升高,IFN-γ、IL-2水平降低,說明Th1/Th2隨治療時間而升高,此結果與Memisx等[17]的研究成果一致。本研究結果表明,右美托咪定干預治療可使感染性休克大鼠Th1/Th2升高。右美托咪定干預使感染性休克大鼠脾臟及胸腺caspase-3、caspase-8、caspase-9表達及淋巴細胞凋亡的陽性細胞數顯著減少,且caspase-9表達的陽性細胞數減少幅度最為明顯。因此,右美托咪定可能通過調節感染性休克大鼠機體內的炎性反應,促進Th1/Th2平衡,減少淋巴細胞凋亡;而且還可能通過調節其體內炎性反應抑制脾臟及胸腺的caspase-3、caspase-8、caspase-9活性,從而也減少淋巴細胞凋亡。
綜上所述,右美托咪定對感染性休克大鼠具有減輕炎癥反應和改善免疫能力的作用,有利于改善預后。
參考文獻:
[1] 薛娜,梁輝,姚輝,等.脾臟在電刺激迷走神經抗大鼠感染性休克中的作用[J].中國危重病急救醫學,2011,23(5):263-276.
[2] 吳云,柯建娟,張宗澤.布托啡諾對感染性休克大鼠細胞因子的影響[J].中華急救醫學雜志,2010,19(2):132-134.
[3] Nance DM,Sanders VM.Autonomic innervation and regulation of the immune system[J].Brain Behav Immun,2011,21(17):736-745.
[4] 王樂,陳磊,李雪芬,等.三氧化二砷對肝癌細胞系SMMC-7721凋亡及Smac caspase-9,caspase-3蛋白表達的影響[J].現代生物醫學進展,2013,13(33):6440-6444.
[5] 章莉莉,孫志堅,張東軍,等.右美托咪定鎮靜對機械通氣膿毒癥休克患者細胞免疫和炎性因子的影響[J].河北醫科大學學報,2013,34(1):33-36.
[6] 張昌鋒,吳秀娟,蔣宗明,等.右美托咪定鎮靜對機械通氣膿毒癥休克患者細胞免疫和炎性因子的影響[J].河北醫科大學學報,2013,34(1):33-36.
[7] 蔣紅濤,陳艷,唐貴文.曼氏裂頭蝴感染小鼠外周血T淋巴細胞亞群及IL-4,IL-10,IFN-γ,TNF-α的動態變化[J].中國病原生物學雜志,2009,4(11):840-843.
[8] Nikonenko BV,Portopopova M,Samala R,et al.Drug therapy of experimental tuberculosis(TB):improved outcome by combining SQl09,a new dimaine antibiotic,with existing TB drugs[J].Antimierob Agents Chemother,2007,51(4):l563-1565.
[9] Boyman O,Surh CD,Sprent J.Potentialuse of IL-2/anti-IL-2 antibody immune complexes for the treatment of cancer and autoimmune disease[J].Expert Opin Biol Ther,2006,6(12):1323-1331.
[10]Foitzik T,Hotz HG,Eibl G et al.Therapy for microcirculatory disorders in severe acute pancreatitis:effectiveness of platelet-activating factor receptor blockade vs.endothelin receptor blockade[J].J Gastrointest Surg,1999,3(3):244-251.
[11]Li B,Tournier C,Davis RJ,et al.Regulation of IL-4 expression by the transcirption factor Jun B duirng T helper cell differentiation[J].EMBOJ,2009,18(2):420-432.
[12]李旭芳,劉玲玲,張菊,等.急性MCMV感染對小鼠脾Thl/Th2/Th17細胞亞群分化的影響[J].中國免疫學雜志,2013,29(5):464-467.
[13]吳文秀,崔利娟,陳煌,等.截斷逆挽方對慢加急性肝衰竭大鼠肝細胞凋亡指數及caspase8,caspase3表達的影響[J].北京中醫藥,2012,31(1):62-64.
[14]Zhuge J,Cederbaum AI.Inhibition of the mitochondrial permeability transition by cyclosporin A prevents pyrazole plus lipopolysaccharide-induced liver injury in mice[J].Free Radic Biol Med,2009,46(3):406-413.
[15]甘建輝,史金麟,胡萬寧,等.右美托咪定在乳腺癌前哨淋巴結活檢術中的臨床應用[J].中國藥房,2013,21(10):921-924.
[16]李志鵬,柳垂亮.右美托咪定的臨床應用進展[J].實用醫學雜志,2013,29(19):3254-3256.
[17]Memisx D,Hekimoglu S,Vatan I,et al.Effects of midazolam and dexmedetomidine on inflammatory responses and gastric intramucosal pH to sepsis,in critically ill patients[J].Br J Anaesth,2007,98(12):550-552.
