上/下調miR-21對結腸癌細胞的生物學作用及對西妥昔單抗藥物敏感性的影響*
2014-08-08 07:24:18李東風謝子鈞段伊帆李子俊
中國病理生理雜志 2014年4期
關鍵詞:結腸癌
鞏 波, 李東風, 謝子鈞, 段伊帆, 李子俊△
(1南方醫科大學,廣東 廣州 510515;廣東省人民醫院, 廣東省醫學科學院 2消化科, 3醫學研究中心,廣東 廣州 510080)
在美國和大部分歐洲國家,結直腸癌是第3位常見的惡性腫瘤,與腫瘤相關的死亡原因排第2位,在這些病人中,大約有20%的病人在獲得臨床診斷時已發生轉移[1]。西妥昔單抗靶向表皮生長因子受體(epidermal growth factor receptor, EGFR),抑制細胞的生長、增殖和轉移,已被美國FDA批準用于KRAS野生型結直腸癌的治療[2]。但是并非所有KRAS野生型結直腸癌都能從西妥昔單抗治療中獲益,據報道在占結直腸癌60%~70% 的KRAS野生型中,只有其中60%獲得治療反應,余下40% KRAS野生型結直腸癌治療無效或耐藥[3]。而迄今研究表明miR-21在人體31種腫瘤中表達異常,也是最常見的異常microRNA之一,是一種致癌性microRNA, 在多種腫瘤的發生和發展中起著重要的作用。本實驗通過體外實驗上調/下調miR-21,探討其對人結腸癌細胞RKO的增殖、克隆形成能力及對西妥昔單抗敏感性的影響,為下一步體內實驗和臨床研究提供依據。
材 料 和 方 法
1 細胞
人結腸癌細胞株RKO(購自中國科學院上海細胞研究所)使用MEM Alpha培養基(Life Technologies)加10%胎牛血清(Life Technologies),5% CO2、37 ℃培養。
2 方法
2.1慢病毒載體的構建 根據人miR-21的基因序列,制備上調/下調miR-21雙鏈DNA Oligo,設計包含目的基因序列和酶切位點的互補序列,LV-miR-21上游引物5’-CGGCCGCGACTCTAGTTATCAAATCCTGCCTGACTG-3’,下游引物5’- ATAAGCTTGATATCG-TCCTCCCTCCATACTGCTG-3’;LV-anti-miR-21上游引物5’-CCGGTCAACATCAGTCTGATAAGCTATTTTTG-3’,下游引物5’-AATTCAAAAATAGCTTATCAGACTGATGTTGA-3’。將引物退火形成帶黏性末端的雙鏈DNA,利用T4 DNA ligase分別連接到GV217和GV159載體上,轉化感受態細菌,挑選轉化的菌落,用菌落PCR進行鑒定和測序。
2.2慢病毒載體的包裝 取生長狀態良好的293T細胞消化重懸后按1∶5比例傳至poly-(D-lysine)(PDL)包被的10 cm皿中,加完全培養基37 ℃、5% CO2細胞培養箱中繼續培養約24 h,細胞密度達50%~70%時即可用于轉染。取1.5 mL/dish Opti-MEM、表達質粒6 μg/dish、包裝質粒Packaging Mix 6 μg/dish,混勻,室溫下溫育5 min;取1.5 mL/dish Opti-MEM和POLOdeli-vererTM3000 Transfection reagent 24 μL/dish,輕輕混勻,室溫下溫育5 min。將兩者混合,室溫下孵育20 min。將DNA與POLOdelivererTM3000 Transfection reagent 的復合液均勻地滴加在293T細胞的培養液中混勻,于37 ℃、5% CO2細胞培養箱中培養,48 h后熒光顯微鏡觀察轉染效率,收集293T細胞上清液,4 ℃暫存;細胞更換新鮮完全培養基后繼續培養至72 h,收集病毒上清,與第1次收集的混合,于 4 ℃、3 000 r/min 離心10 min,用0.45 μm 醋酸纖維素膜的濾器過濾于Beckman SW28離心管中,每管裝36 mL病毒,4 ℃、22 000 r/min 離心2 h,棄上清,每管用130 μL預冷的PBS溶解沉淀,每管100 μL分裝,-80 ℃保存。
2.3慢病毒滴度測定 測定前1 d,取測定滴度所需的293T細胞,每孔加100 μL,4×104個細胞接種到96孔板,根據病毒的預期滴度,準備7~10個無菌的EP 管,在每個管中加入 90 μL 的無血清培養基,將待測定的病毒原液 10 μL 加入到第1個管中,混勻后,取 10 μL 加入到第2個管中,繼續相同的操作直到最后1管。選取所需的細胞孔,吸去 90 μL 培養基,丟棄,加入 90 μL 稀釋好的病毒溶液,放入培養箱培養,24 h后,加入完全培養基 100 μL。4 d 后,觀察熒光表達情況。病毒滴度=陽性細胞數×計量孔稀釋倍數/接種病毒體積。
2.4細胞感染 取生長狀態良好的RKO細胞,胰酶消化后用全培養基重懸細胞,每孔2×105個細胞接種到6孔板,37 ℃、5% CO2細胞培養箱中繼續培養約24 h。取6孔板中的一孔細胞進行消化,計數細胞,根據MOI和細胞數量計算所需要的病毒量稀釋到培養基中;吸去舊培養基,加入含病毒液的培養基,同時設置陰性對照空載體組和空白對照組,12 h后換液,繼續培養72 h后在熒光顯微鏡下觀察熒光表達情況,初步估計慢病毒感染效率。
2.5qRT-PCR檢測miR-21的表達 收集上調/下調miR-21、陰性對照及空白對照的RKO細胞,采用Trizol法提取細胞總RNA并測定總RNA的濃度及純度。根據逆轉錄試劑盒ReverTra Ace qPCR RT Kit(Toyobo)合成cDNA,反應條件為37 ℃ 15 min,98 ℃ 5 min,4 ℃終止。取逆轉錄產物作為模板,用Realtime PCR Master Mix(SYBR Green)試劑盒(Toyobo)進行qRT-PCR檢測,以U6為內參照,反應條件為95 ℃ 30 s, 然后95 ℃ 10 s, 60 ℃ 20 s,72 ℃ 30 s,45個循環,每個樣品設置4個復孔。采用Lightcycle羅式熒光定量PCR儀進行檢測。采用2-ΔΔCt法對數據進行分析。
2.6細胞增殖實驗 取對數生長期的上調/下調miR-21、陰性對照及空白對照的RKO細胞,胰酶消化,細胞計數后按照每孔1×104個細胞接種到96孔板,每組設置6個復孔,與實驗孔平行設不加細胞只加培養液的空白對照孔,分別接種4塊板,于培養后24 h、48 h、72 h和96 h分別加入MTT溶液(5 g/L)20 μL/well,37 ℃繼續孵育4 h,終止培養,小心吸棄孔內培養上清液。每孔加入150 μL DMSO,振蕩10 min,使結晶物充分溶解;于490 nm波長處測定A值。
2.7克隆形成實驗 配置1.2%和0.7%無菌低熔點瓊脂液體,40 ℃維持,使之保持溶解狀態。按1∶1混合1.2%瓊脂糖和2×完全培養基,按每孔取2 mL混合液注入6孔板中,待冷卻凝固,備用。收集對數生長期的上調/下調miR-21、陰性對照及空白對照的RKO細胞,制成1×107/L的細胞懸液;按1∶1比例將0.7%瓊脂和2×完全培養基在無菌試管中混合后,取1 mL混合液+0.1 mL單細胞懸液(1×103cells/well),充分混勻、注入已鋪有底層瓊脂的培養板中,在室溫放置20 min使細胞瓊脂懸液凝固。將6孔板置于CO2培養箱中37 ℃進行培養。培養10 d,觀察細胞克隆形成情況并計數。
2.8凋亡實驗 胰酶消化上調/下調miR-21、陰性對照及空白對照的RKO細胞,分別收集(1~5)×105細胞。在50 μL binding buffer中加入5 μL 7-AAD染液,混勻;收集細胞中加入上述7-AAD染液,混勻,室溫避光反應5~15 min;再加入450 μL binding buffer混勻后加入1 μL Annexin V-PE混勻,室溫避光反應5~15 min;1 h內流式細胞術檢測。
2.9西妥昔單抗敏感性檢測 取對數生長期的上調/下調miR-21、陰性對照及空白對照的RKO細胞,胰酶消化,細胞計數后按照每孔1×104個細胞接種到96孔板,24 h后,加入300 mg/L的西妥昔單抗,不同細胞設3復孔,繼續培養24 h后,棄培養液,每孔加入100 μL無血清培養基及10 μL CCK-8,繼續培養,每30 min在波長450 nm測定吸光度A值,直至未加藥孔的A值為1.0~1.2時為最佳時間。細胞抑制率=(實驗孔A值-空白孔A值)/(未加藥孔A值-空白孔A值)×100%。
3 統計學處理
使用SPSS 13.0 軟件分析。數據以均數±標準差(mean±SD)表示,采用單因素方差分析或重復測量資料的方差分析比較轉染不同慢病毒的細胞間miR-21的表達、細胞增殖能力、克隆形成能力、細胞凋亡、藥物敏感性等的差異,以P<0.05為差異有統計學意義。
結 果
1 重組慢病毒的包裝與轉染
將構建好的慢病毒干擾質粒轉染到293T細胞中,包裝產生重組慢病毒顆粒,通過觀察綠色熒光,可見90%以上的細胞帶有綠色熒光,見圖1。收集濃縮病毒顆粒后測定LV-miR-21與LV-anti-miR-21的滴度分別為3.0×1012TU/L和2.0×1012TU/L,將病毒顆粒分別感染人結腸癌RKO細胞,觀察綠色熒光,感染效率在80%以上,見圖2。

Figure 1. 293T cells transfected with the lentivirus interference plasmids(×100). A,B: LV-miR-21; C,D: LV-anti-miR-21.A,C:fluorescence view; B,D: phase-contrast view.

Figure 2. RKO cells transfected with different lentiviral vectors(×100). A1, A2: LV-miR-21; B1, B2: LV-miR-21 NC; C1,C2: LV-anti-miR-21; D1, D2: LV-anti-miR-21 NC.A1,B1,C1,D1: fluorescence view; A2,B2,C2,D2: phase-contrast view.
2 qRT-PCR檢測結腸癌細胞RKO轉染慢病毒干擾載體后miR-21的表達
轉染LV-miR-21、LV-anti-miR-21、相應陰性對照載體和空白組的miR-21相對表達量分別為9.45±0.56、0.55±0.13、1.17±0.10、1.11±0.09和1.00±0.00,其中LV-miR-21組的表達量高于其陰性對照組及空白組,差異有統計學意義(P<0.05);LV-anti-miR-21組的表達量低于其陰性對照組及空白組,差異有統計學意義(P<0.05),而陰性對照組與空白組的表達量之間無明顯差異(P>0.05),見圖3。
3 MTT檢測人結腸癌細胞轉染不同慢病毒后細胞增殖能力的變化
MTT實驗結果表明轉染LV-anti-miR-21后,隨著時間的增長,細胞的增殖能力明顯降低;轉染LV-miR-21后,隨著時間的增長,細胞的增殖能力明顯增強。在第2天、3天和4天,LV-miR-21組、LV-anti-miR-21組和空白組之間細胞增殖能力有明顯差異(P<0.01),見圖4。
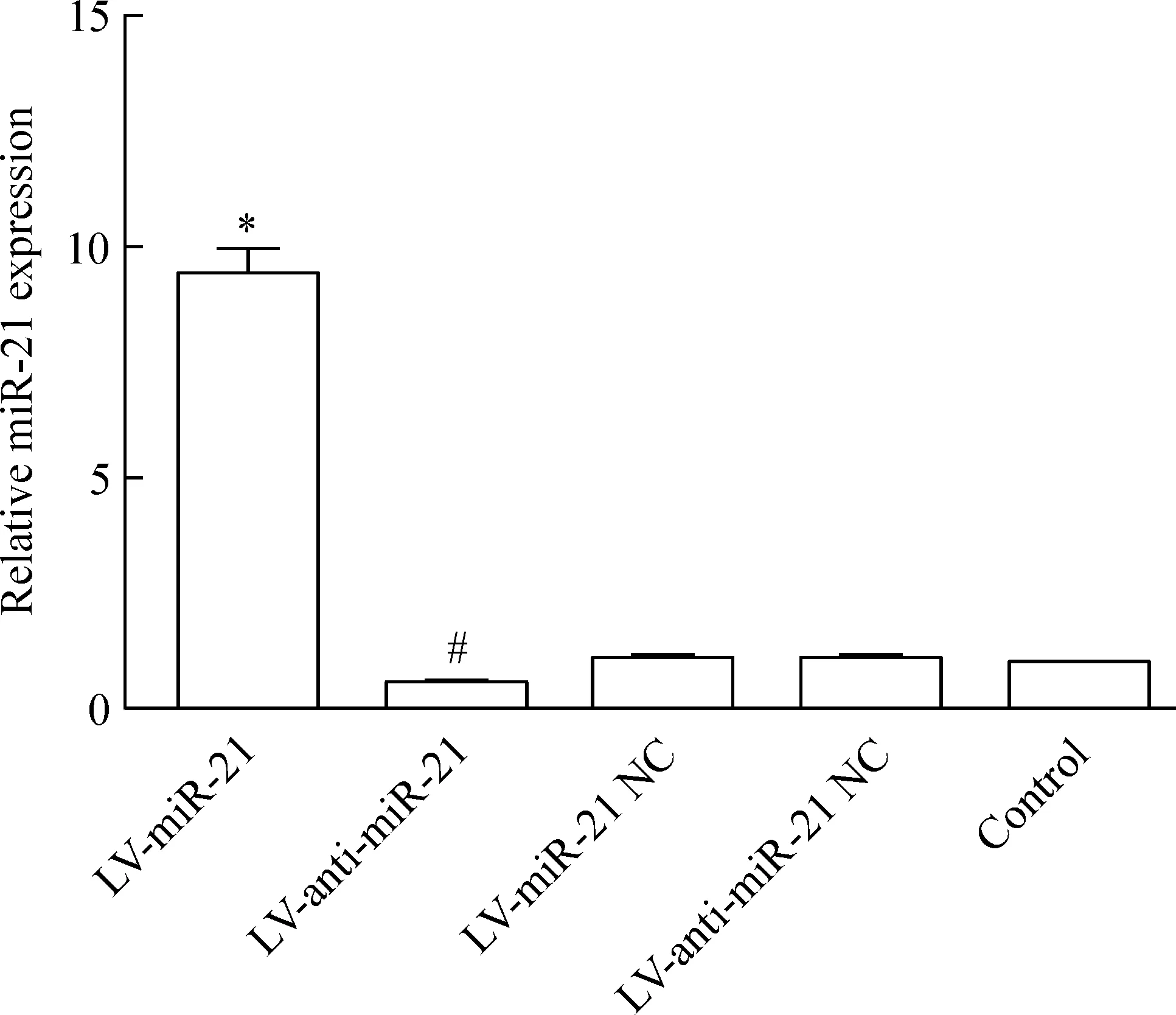
Figure 3. Expression of miR-21 in transfected RKO cells.Mean±SD.n=4. *P<0.05 vs LV-anti-miR-21, LV-miR-21 NC or control; #P<0.05 vs LV-miR-21 or LV-anti-miR-21 NC.
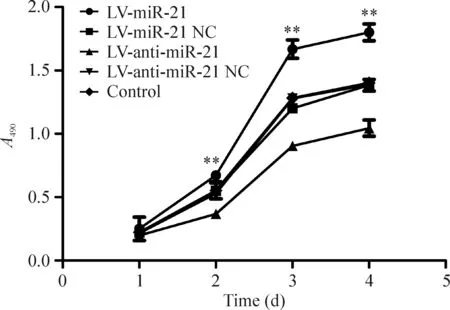
Figure 4. Proliferation of RKO cells with different treatments.Mean±SD.n=6.**P<0.01 vs LV-anti-miR-21.
4 轉染不同慢病毒后克隆形成能力的變化
LV-miR-21組的細胞克隆形成能力明顯高于其陰性對照組、空白組和LV-anti-miR-21組(P<0.01);而LV-anti-miR-21組的細胞克隆形成能力低于其陰性對照組和空白組(P<0.01),見圖5。
5 轉染不同慢病毒后細胞凋亡的變化
LV-anti-miR-21組的早期凋亡率明顯高于LV-miR-21組(P<0.01),見圖6。這說明下調miR-21可以促進細胞的凋亡。
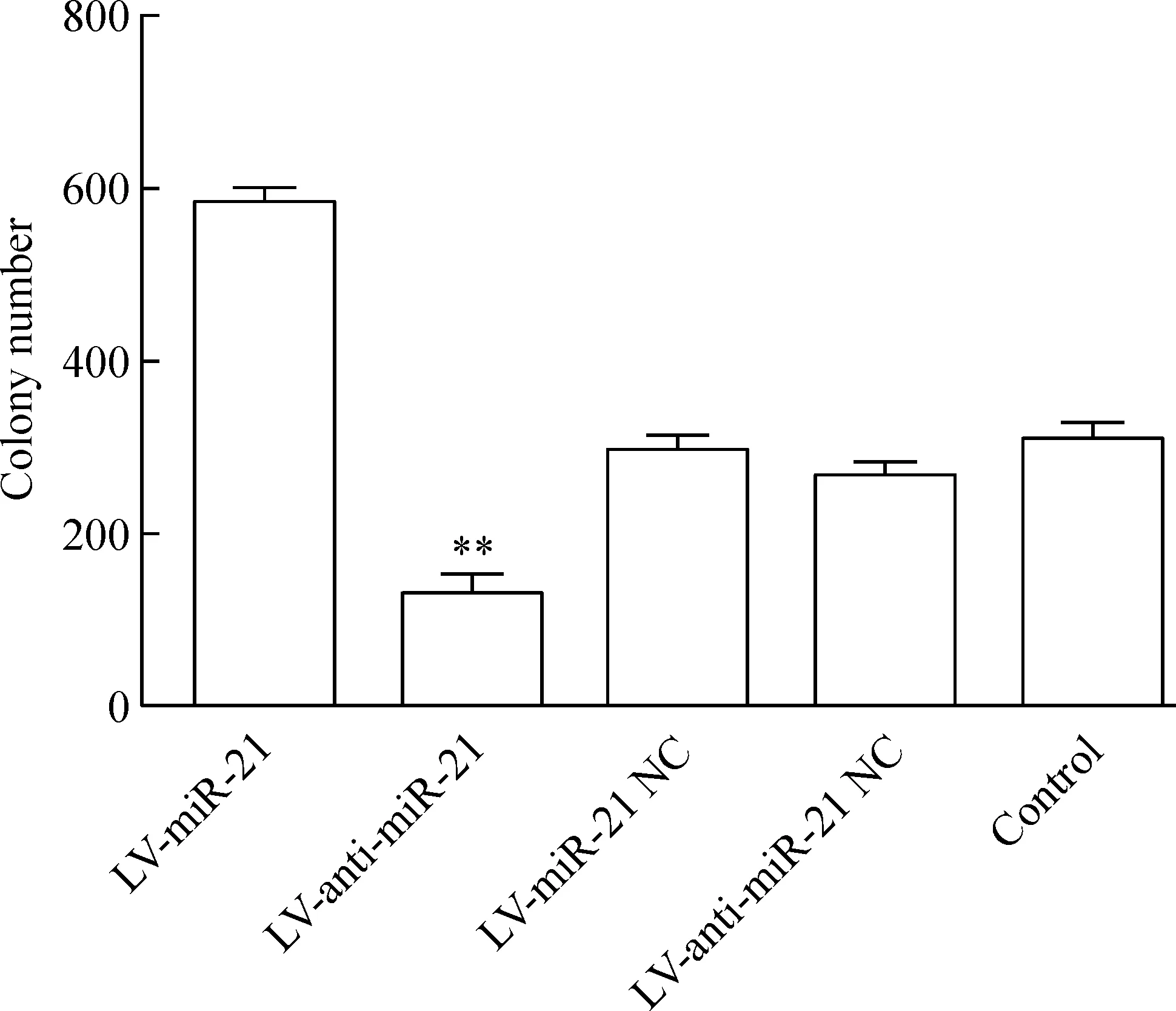
Figure 5. Colony-forming ability of the RKO cells with different treatments.Mean±SD.n=3.**P<0.01 vs LV-miR-21.
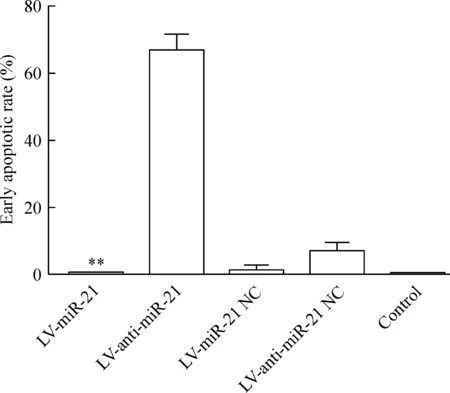
Figure 6. The early apoptotic rate of the RKO cells with different treatments.Mean±SD.n=3.**P<0.01 vs LV-anti-miR-21.
6 下調miR-21可增強腫瘤細胞對靶向治療藥物西妥昔單抗的敏感性
在300 mg/L西妥昔單抗作用下,LV-anti-miR-21組的細胞抑制率高于LV-miR-21組,差異有統計學意義(P<0.05),見圖7。
討 論
MicroRNA是一類內源性非編碼小分子單鏈RNA,能夠調控mRNA的表達或翻譯, 在生物體的發育、增殖、分化及凋亡等方面發揮重要的生理作用[4]。近年來, 越來越多的研究表明, 許多micro-RNA可作為原癌基因或者抑癌基因, 在腫瘤的發生和發展中扮演著重要的角色。目前發現,食管癌、胃癌、肝癌、胰腺癌、膽管癌、結直腸癌等腫瘤中miR-21的水平明顯上調。miR-21在眾多腫瘤中呈現出癌基因樣的作用,在腫瘤的發生發展、增殖、凋亡、侵襲、轉移等多種生物學行為中發揮著重要的作用。在結腸癌的研究中,Schetter等[5]檢測了84 例結腸癌組織及其癌旁正常組織的microRNA表達譜,發現有37個microRNA表達存在差異, 其中miR-21在癌組織中表達顯著上調, 證實miR-21與結腸癌的發生密切相關。也有研究發現miR-21在乳腺癌組織中表達顯著上調,其表達上調水平與乳腺癌臨床分期、淋巴結轉移及預后相關[6]。許多研究表明,microRNA具有調節下游靶基因表達的作用。在大多腫瘤中高表達的miR-21很可能是通過調節增殖、凋亡相關基因的表達而發揮促進腫瘤發生發展的作用。Park等[7]研究發現,低濃度的miR-21反義引物的核苷酸顯著抑制胰腺癌細胞株的增殖,抑制miR-21表達可使胰腺癌細胞株HS766T的凋亡增加3~6倍。我們的研究表明下調miR-21能夠明顯抑制結腸癌細胞對增殖及克隆形成能力等。

Figure 7. The inhibitory rate of cetuximab (300 mg/L) on the RKO cells with different treatments.Mean±SD.n=3. *P<0.05 vs LV-anti-miR-21.
此外,miR-21在腫瘤藥物的耐藥性方面也有重要作用[8]。目前研究認為miR-21調控腫瘤細胞對多種藥物敏感性的機制主要與miR-21參與調控細胞周期、抑制細胞凋亡有關。最初由Meng等[9]關于microRNA與化療藥物敏感性的研究發現,抑制miR-21的表達能夠增加膽管癌細胞對吉西他濱的敏感性。深入研究機制,還發現miR-21通過激活PTEN依賴的PI3K信號通路從而調節吉西他濱介導的細胞凋亡,說明miR-21在調節腫瘤細胞化療敏感性方面有重要作用。Park等[7]也發現反義miR-21能夠增強吉西他濱的敏感性。我們通過上調/下調人結腸癌細胞RKO的miR-21表達水平,分析了miR-21對RKO細胞增殖、凋亡、克隆形成能力及對西妥昔單抗敏感性的影響,發現下調miR-21能夠抑制腫瘤細胞的增殖及克隆,增加腫瘤細胞對西妥昔單抗藥物的敏感性。這些結果表明miR-21的高表達可作為結直腸癌的分子標記物,下調miR-21的表達,在結直腸癌等腫瘤治療中有應用前景,值得深入研究和驗證。
[參 考 文 獻]
[1] Lamas MJ, Duran G, Gallardo E. Anti-EGFR therapy in first-line colorectal cancer[J]. Expert Rev Anticancer Ther, 2011, 11(10):1499-1503.
[2] Adelstein BA, Dobbins TA, Harris CA, et al. A systematic review and meta-analysis of KRAS status as the determinant of response to anti-EGFR antibodies and the impact of partner chemotherapy in metastatic colorectal cancer[J]. Eur J Cancer, 2011, 47(9):1343-1354.
[3] García-Sáenz JA, Sastre J, Díaz-Rubio García E. Biomarkers and anti-EGFR therapies for KRAS wild-type metastatic colorectal cancer[J]. Clin Transl Oncol, 2009, 11(11):737-747.
[4] Fujita S, Ito T, Mizutani T, et al. miR-21 gene expression triggered by AP-1 is sustained through a double-negative feedback mechanism[J]. J Mol Biol, 2008, 378(3):492-504.
[5] Schetter AJ, Leung SY, Sohn JJ, et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma[J]. JAMA, 2008, 299(4):425-436.
[6] 顏黎栩,黃馬燕,吳秋良,等. miR-21表達異常與乳腺癌臨床病理特征及預后的關系 [J]. 中國病理生理雜志, 2009, 25( 4): 676- 681.
[7] Park JK, Lee EJ, Esau C, et al. Antisense inhibition of microRNA-21 or -221 arrests cell cycle, induces apoptosis, and sensitizes the effects of gemcitabine in pancreatic adenocarcinoma[J]. Pancreas, 2009, 38(7):e190-e199.
[8] Meng F, Henson R, Lang M, et al. Involvement of human micro-RNA in growth and response to chemotherapy in human cholangiocarcinoma cell lines[J]. Gastroenterology, 2006, 130(7):2113-2129.
[9] Meng F, Henson R, Wehbe-Janek H, et al. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer[J]. Gastroenterology, 2007, 133 (2):647-658.
猜你喜歡
天津醫科大學學報(2019年6期)2019-08-13 07:04:32
西南國防醫藥(2016年7期)2016-12-01 06:01:15
腹腔鏡外科雜志(2016年10期)2016-06-01 12:10:08
中國衛生標準管理(2015年6期)2016-01-14 05:17:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
中國當代醫藥(2015年26期)2015-03-01 02:06:57
西南軍醫(2015年6期)2015-01-23 01:25:50
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
中華介入放射學電子雜志(2014年1期)2014-02-02 05:24:06
