2010-2011年長江河口及鄰近水域春夏季仔稚魚群落結(jié)構(gòu)及其多樣性的研究
2014-06-01 12:30:10蔣玫李磊沈新強(qiáng)全為民
海洋學(xué)報(bào) 2014年6期
關(guān)鍵詞:優(yōu)勢(shì)
蔣玫,李磊,沈新強(qiáng),全為民
(1.中國水產(chǎn)科學(xué)研究院東海水產(chǎn)研究所,上海 200090)
2010-2011年長江河口及鄰近水域春夏季仔稚魚群落結(jié)構(gòu)及其多樣性的研究
蔣玫1,李磊1,沈新強(qiáng)1,全為民1
(1.中國水產(chǎn)科學(xué)研究院東海水產(chǎn)研究所,上海 200090)
根據(jù)2010—2011年春夏季對(duì)長江河口及鄰近水域進(jìn)行了仔稚魚生態(tài)調(diào)查,應(yīng)用單元和多元統(tǒng)計(jì)方法分析了春夏季長江河口及鄰近水域仔稚魚群落結(jié)構(gòu)。4個(gè)航次調(diào)查共獲得27個(gè)種類的仔稚魚,隸屬5目14科。優(yōu)勢(shì)種類主要包括鳀Engraulis japonicus、鳳鱭Coilia mystus、康氏小公魚Stolephorus commersonii、鮻Liza haematocheila、黃姑魚Nibea albiflora、黃鰭刺鰕虎魚Acanthogobius flavimanus和寡鱗飄魚Pseudolaubuca engraulis等。Shannon-Weaver指數(shù)(H′)在調(diào)查周期內(nèi)的波動(dòng)幅度較大,且2011年明顯低于2010年。仔稚魚群落結(jié)構(gòu)和多樣性指數(shù)春季年間較夏季年間差異性顯著。調(diào)查區(qū)夏季群落結(jié)構(gòu)年間相似性不高,春季群落結(jié)構(gòu)格局年際變化則較明顯。聚類分析表明,春夏季長江河口近岸水域仔稚魚群落可劃分為2個(gè)站位組,分別對(duì)應(yīng)于長江河口淡水水域和長江口咸水水域。淡水水域組群落和咸水水域組群落總豐度與鹽度相關(guān)性較高。表明鹽度可能是造成長江河口仔稚魚群落結(jié)構(gòu)時(shí)空差異性的主要原因。
長江口;仔稚魚;群落結(jié)構(gòu);聚類分析
1 引言
河口海區(qū)以其獨(dú)特的地理位置、水動(dòng)力條件和物質(zhì)基礎(chǔ),為眾多海洋生物的生存和繁衍提供了有利的生存條件,導(dǎo)致許多魚類的幼體階段均生活在這一生態(tài)系統(tǒng)中,完成其補(bǔ)充過程,從而使河口海區(qū)成為大量海洋魚類重要的產(chǎn)卵、育肥場,孕育了豐富的魚類資源[1—2]。長江口海區(qū)因這種特殊的生態(tài)特點(diǎn),注定了該區(qū)域魚類補(bǔ)充群體——魚類浮游生物種類組成的復(fù)雜性和多樣性。羅秉征等曾捕獲魚卵90萬個(gè),仔稚魚7萬尾,隸屬54科、100種,是該區(qū)域有記錄以來最多的一次,具有很高的多樣性[3]。隨后也有不少學(xué)者相繼調(diào)查和分析了長江口水域的魚卵和仔稚魚組成,但種類數(shù)量明顯減少[4—6]。河口魚類浮游生物群落結(jié)構(gòu)及其季節(jié)變化格局,既受制于生殖種群的資源量,也與水域環(huán)境的穩(wěn)定性密不可分。近年來,由于長江流域經(jīng)濟(jì)快速發(fā)展,人類對(duì)該流域資源與環(huán)境的利用愈來愈多,最終給長江口及鄰近海域的生態(tài)與環(huán)境造成了巨大壓力。魚類浮游生物群落的研究在生態(tài)環(huán)境評(píng)價(jià)和監(jiān)測(cè)及評(píng)價(jià)人類活動(dòng)對(duì)河口影響等方面發(fā)揮著越來越重要的作用[7]。本文根據(jù)2010年至2011年5月和8月4個(gè)季度航次最新調(diào)查資料,探討長江口及其鄰近水域仔稚魚群落物種群落結(jié)構(gòu)和多樣性,旨在為該水域環(huán)境的評(píng)估和監(jiān)測(cè)提供基礎(chǔ)資料。
2 材料與方法
分別于2010年的5月(春季)和8月(夏季)和2011年的5月(春季)和8月(夏季),在長江口海域(30°48.866′~31°46.447′N,121°0.112′~122°36.431′E)設(shè)置30個(gè)調(diào)查站位(圖1)。仔稚魚樣品采集參照《海洋調(diào)查規(guī)范》,采用淺水I型浮游生物拖網(wǎng)(網(wǎng)長145 cm,網(wǎng)口內(nèi)徑50 cm,網(wǎng)口面積0.2 m2),水平拖網(wǎng)10 min,拖速約2.0 n mile/h;樣品保存于5%的福爾馬林海水混合溶液中。在實(shí)驗(yàn)室內(nèi)從浮游生物樣品中挑取仔稚魚標(biāo)本,采用生物解剖鏡,對(duì)各站標(biāo)本進(jìn)行種類鑒定、個(gè)體計(jì)數(shù)和發(fā)育階段的判別。水平網(wǎng)以每網(wǎng)全網(wǎng)的實(shí)際數(shù)量(尾)為指標(biāo)來計(jì)算仔稚魚的總豐度(尾/網(wǎng))。種類的鑒定按照《中國近海魚卵與仔魚》[8]和《上海魚類志》[9],同時(shí)比照歷史樣品中的模式標(biāo)本。利用多參數(shù)水質(zhì)分析儀(YSI-600)進(jìn)行溫度、鹽度等環(huán)境因子的測(cè)定。泥沙和徑流量大小根據(jù)《2010年和2011年中國河流泥沙公報(bào)》提供的數(shù)據(jù)做參考[10—11]。
運(yùn)用多樣性指數(shù)法,分析各航次種類組成的多樣性[]。
Margalef豐富度指數(shù):
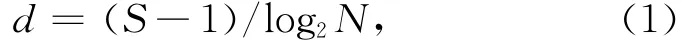
式中,S為總種數(shù),N為總個(gè)體數(shù)。
Shannon-Weaver多樣性指數(shù):
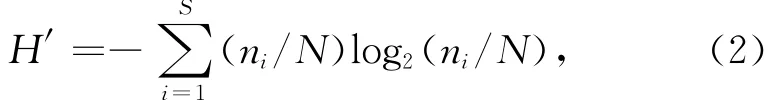
式中,H′為種類多樣度,ni為第i種魚的個(gè)體數(shù),N為魚類總個(gè)體數(shù)。
Pielou種類均勻度:

式中,J為魚類種類均勻度,S為魚類種類總個(gè)體數(shù)。
運(yùn)用相對(duì)重要性指數(shù)IRI(the index of relative importance)對(duì)仔稚魚種類的優(yōu)勢(shì)度進(jìn)行分析[13]。
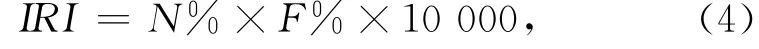
式中,N%為個(gè)體數(shù)量百分比;F%為頻次比例。本文選取IRI值大于100的種類為優(yōu)勢(shì)種,IRI值大于10的種類為常見種,小于10的為稀有種。
不同年份間仔稚魚群落結(jié)構(gòu)組成的差異采用Bray-Curtis相似性指數(shù)來分析,應(yīng)用CLUSTER聚類分析來研究群落結(jié)構(gòu)[14],單因子相似性分析(ANOSIM)用于不同組群落結(jié)構(gòu)差異的顯著性檢驗(yàn)。生物-環(huán)境匹配分析(biota-environment matching,BIOENV)使用PRIMER 5.0軟件計(jì)算。采用SPSS 12.0對(duì)不同季節(jié)種類數(shù)和豐度的差異顯著性進(jìn)行單因子方差分析(one-way ANOVA)。
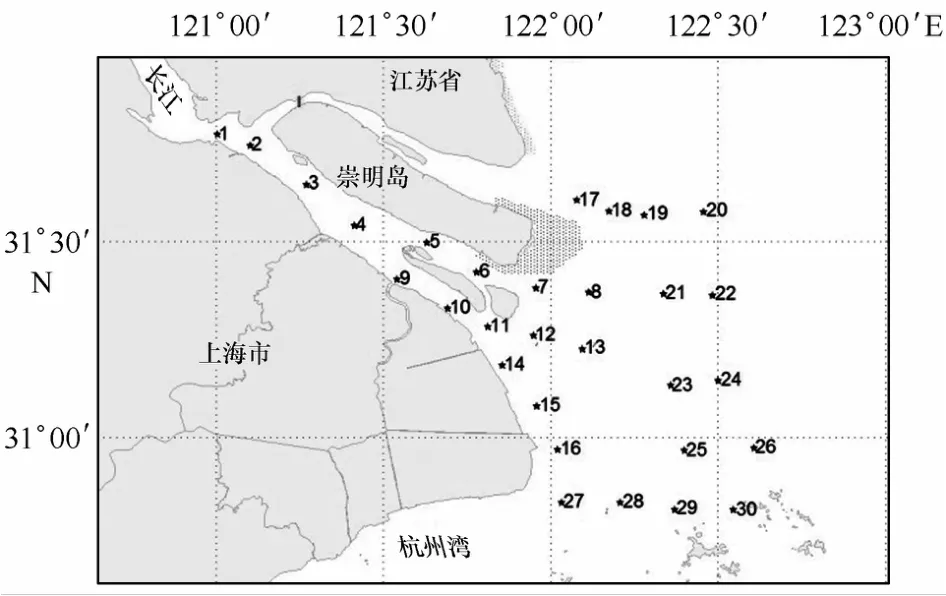
圖1 采樣調(diào)查站位
3 結(jié)果
3.1 仔稚魚豐度和物種組成
調(diào)查期間共獲得仔稚魚5 172尾,27個(gè)種類,隸屬5目14科,其中蝦虎魚科魚類種類最多(5種),占總種數(shù)的18.5%。該水域魚類浮游生物群落由淡水魚類、半咸水魚類和沿岸、近海魚類4種生境類型組成,尤其是河口性和近岸性魚類占絕對(duì)優(yōu)勢(shì)。如2010 年5月總豐度較高主要為鳳鱭Coilia mystus和康氏小公魚Stolephorus commersonii河口性和近岸性魚類,其對(duì)總豐度貢獻(xiàn)最大,兩者占該航次總豐度的94.4%。2010年8月和2011年8月鳀的豐度則占魚類總豐度的64.6%以上。2011年5月總豐度較高的為鮻Liza haematocheila和鳀,占總豐度的78.6%以上。從出現(xiàn)頻率來看,河口性和近岸性魚類(鳀、鳳鱭和康氏小公魚)在4個(gè)航次出現(xiàn)的頻率最高(見表1)。
3.2 優(yōu)勢(shì)種類
依據(jù)IRI指數(shù)確定的優(yōu)勢(shì)種及其組成見表2。2010年5月優(yōu)勢(shì)度較高的種類只有鳳鱭和康氏小公魚;2010年8月,優(yōu)勢(shì)種組成中增加了鳀和淡水種寡鱗飄魚Pseudolaubuca engraulis;2011年5月康氏小公魚和寡鱗飄魚的優(yōu)勢(shì)地位喪失,鮻和黃姑魚成為優(yōu)勢(shì)種。2011年8月的優(yōu)勢(shì)種組成中增加了黃鰭刺鰕虎魚Acanthogobius flavimanus,但減少了鮻和黃姑魚2個(gè)優(yōu)勢(shì)種類,鳀仍舊為第一優(yōu)勢(shì)種。從表2可以看出:4個(gè)航次鳳鱭始終在長江口仔稚魚中占據(jù)優(yōu)勢(shì)地位,且春季的優(yōu)勢(shì)度逐年下降,夏季的優(yōu)勢(shì)度則逐年增加。鳀除在2010年5月出現(xiàn)過波動(dòng)外,在其他時(shí)間均處于絕對(duì)優(yōu)勢(shì)地位;夏季的優(yōu)勢(shì)度明顯要高于春季。康氏小公魚除2011年5月沒有出現(xiàn)外,其他時(shí)間也均處于優(yōu)勢(shì)地位,但優(yōu)勢(shì)度季節(jié)變化規(guī)律性不明顯,春季的優(yōu)勢(shì)度逐年下降,夏季的優(yōu)勢(shì)度逐漸增加。
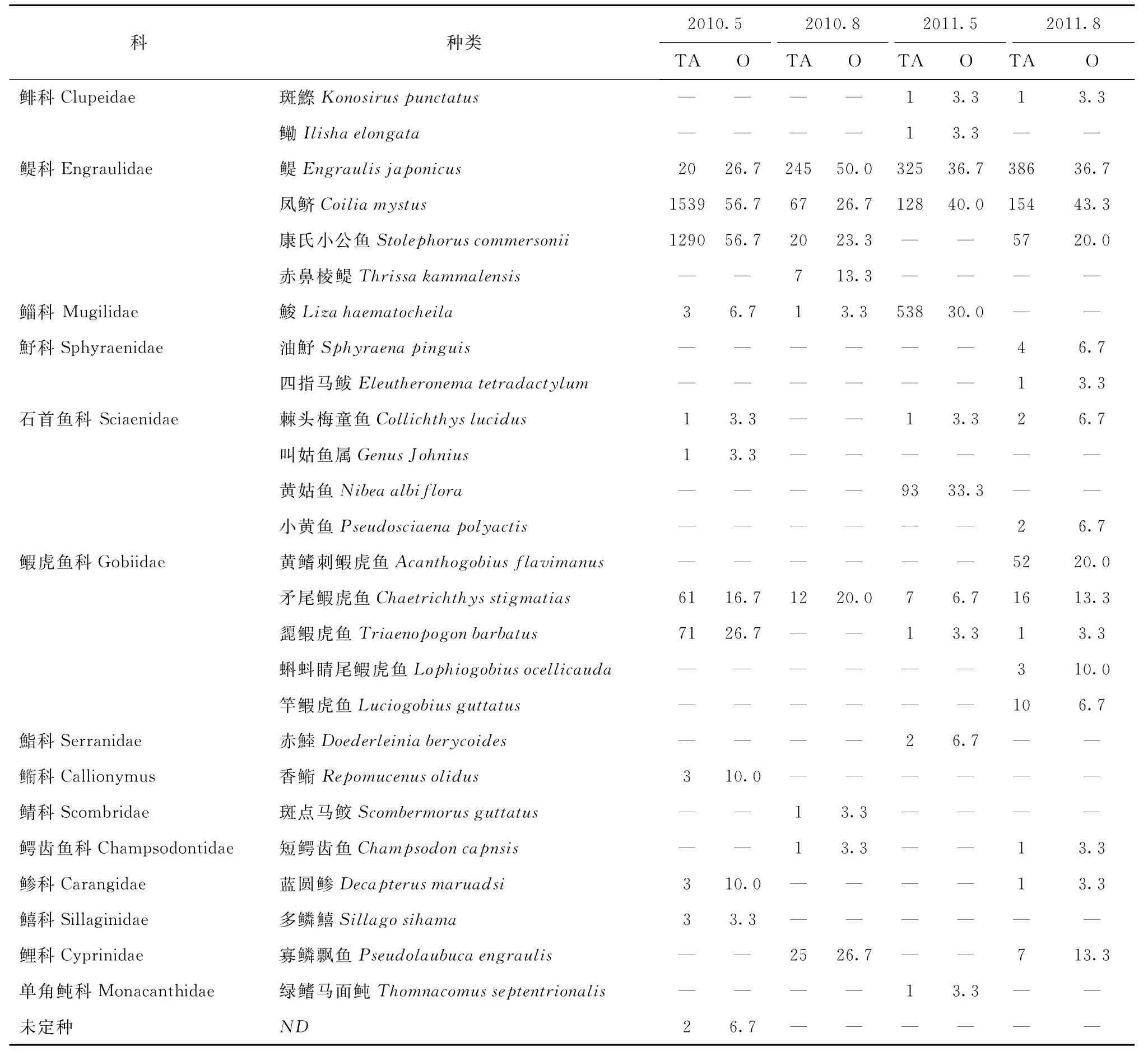
表1 仔稚魚群落的總豐度(TA)和出現(xiàn)頻率(O)
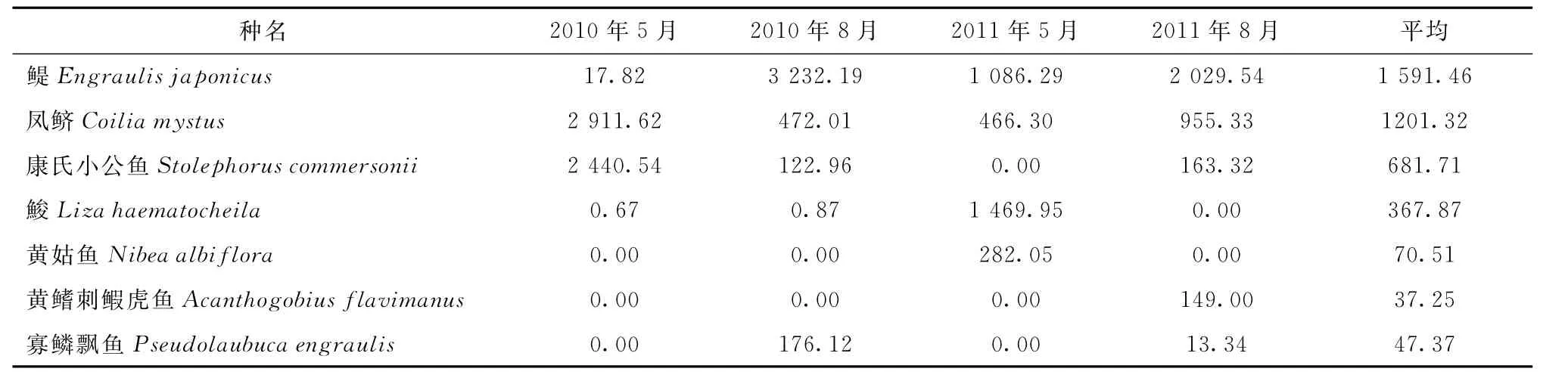
表2 仔稚魚優(yōu)勢(shì)種類組成(IRI指數(shù))
3.3 群落多樣性的時(shí)間動(dòng)態(tài)
在4個(gè)調(diào)查航次中出現(xiàn)的仔稚魚的種類分別為12、9、11和16種,呈現(xiàn)春夏季交錯(cuò)高低變化趨勢(shì)。在研究區(qū)域內(nèi),種類豐富度指數(shù)、均勻度和多樣性指數(shù)變化趨勢(shì)相同,均以2010年5月航次最高,2011 年5月航次最低。對(duì)生物多樣性指數(shù)的ANOVA分析發(fā)現(xiàn),2010年各多樣性指數(shù)季節(jié)性變化較顯著(p<0.05),2011年季節(jié)性差異較小,除了Shannon-Weaver指數(shù)春夏季有顯著差異(p<0.05)外,豐富度和均勻度均無顯著性差異(p>0.05)。從年間變化趨勢(shì)看,春季,Shannon-Weaver指數(shù)和均勻度兩年間存在較顯著差異外(p<0.05),豐富度無顯著差異(p>0.05)。夏季,除豐富度兩年間有較顯著差異外(p<0.05),Shannon-Weaver指數(shù)和均勻度均無顯著差異(p>0.05)。可以看出,2010年較2011年多樣性季節(jié)性變化更顯著,春季年間變化較夏季年間變化更顯著(表3)。

表3 群落多樣性的時(shí)間動(dòng)態(tài)
3.4 群落相似性的時(shí)空變化
計(jì)算出各調(diào)查航次的仔稚魚群落相似性指數(shù)見表4。總體上,各航次的相似性并不高,均在0.6以下。從季節(jié)年間變化看,夏季兩年間的相似性(0.55)較春季(0.50)略高,說明春季年間群落結(jié)構(gòu)上波動(dòng)較大。但兩年的季節(jié)相似性變動(dòng)不大,2010 年5月和8月的相似性為0.38,2011年5月和8月的相似性為0.40,兩者僅相差0.02。從某種意義上說明調(diào)查區(qū)季節(jié)性的群落結(jié)構(gòu)格局變化不明顯。
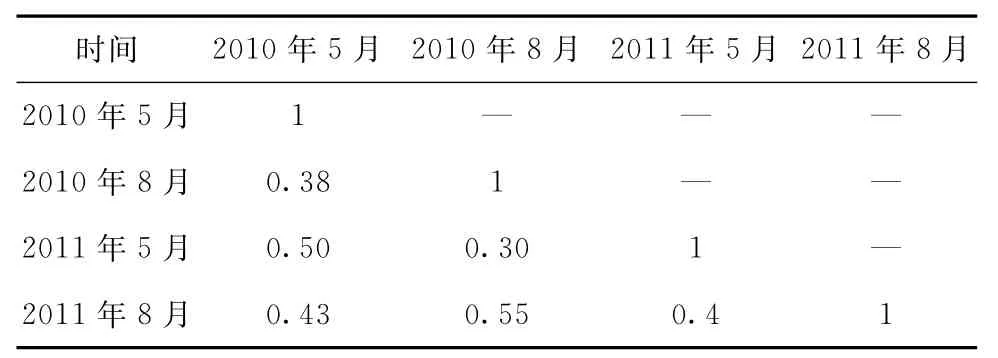
表4 群落相似性的時(shí)間動(dòng)態(tài)
圖2為30個(gè)調(diào)查站位仔稚魚豐度的聚類分析圖,從圖中可以看出,在35.00%的相似性水平上,可以把4個(gè)季度月的樣本分為長江口淡水群落組(A)和長江口咸水群落組(B)2個(gè)區(qū)域,其差別主要與區(qū)域的鹽度有直接關(guān)系。A組內(nèi)包括16個(gè)站位,位于長江口門以內(nèi),主要以長江沖淡水為主,鹽度范圍基本都在1以下,屬于淡水區(qū)。B組內(nèi)包括14個(gè)站位,位于長江口門外側(cè),主要以高鹽外海水為主(鹽度范圍為21.05~35.84)。應(yīng)用ANOSM分析進(jìn)行顯著性檢驗(yàn),物種的檢驗(yàn)結(jié)果為R=0.594(R<1),p<0.05,表明2個(gè)群聚存在顯著性差異。A組中,1號(hào)雖然在長江入海口端,而12號(hào)站和16號(hào)站距離長江入海口較遠(yuǎn),靠近攔門沙處,雖然彼此距離較遠(yuǎn),但依據(jù)聚類分析呈現(xiàn)出較高的相似性,說明群落分布是在部分站位組間呈鑲嵌狀的格局。在B組群落中同樣有這種情況出現(xiàn),例如崇明島北支的18號(hào)和蘆潮港附近海域的27號(hào)站兩者距離遠(yuǎn)相似度卻十分相近。
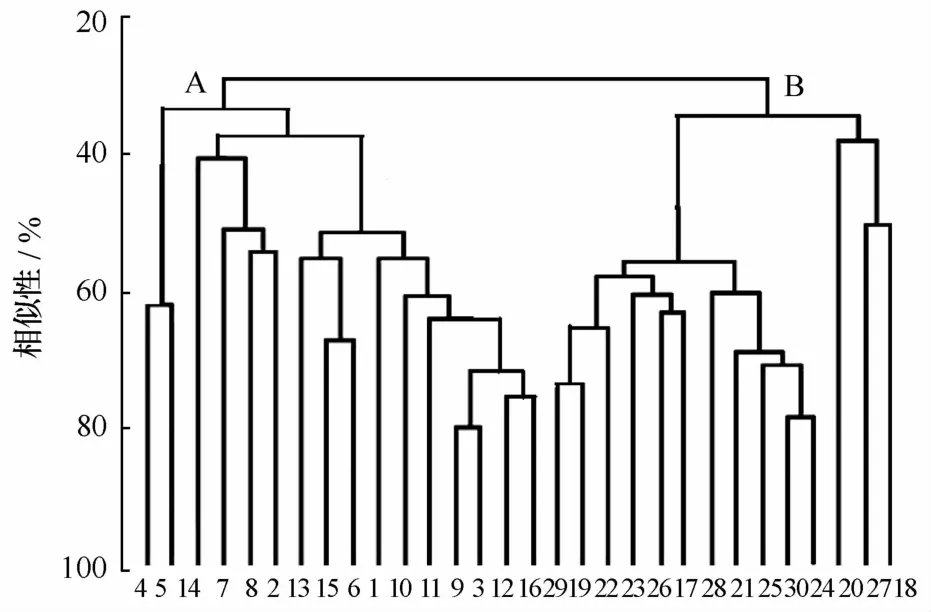
圖2 仔稚魚豐度的不同站位Bray-Curtis聚類分析
3.5 群落結(jié)構(gòu)與環(huán)境因子的關(guān)系
各季節(jié)仔稚魚類群落組成與泥沙、徑流量、水溫、鹽度以及水深等環(huán)境因子的關(guān)系較為復(fù)雜,見表5所示。就A組群落的總豐度與各環(huán)境因子的關(guān)系而言,與徑流量和鹽度的相關(guān)系數(shù)較高(r>0.30),與水深的相關(guān)關(guān)系最低(r<0.20);就B組群落的總豐度與各環(huán)境因子的關(guān)系而言,與鹽度相關(guān)系數(shù)較高(r>0.45),與水深的相關(guān)關(guān)系最低(r<0.30);對(duì)于整個(gè)區(qū)域,與鹽度的相關(guān)關(guān)系較高(r>0.44)。各組群落總豐度與環(huán)境因子特別是鹽度的季節(jié)性和年際間差異性較顯著(p<0.05)。
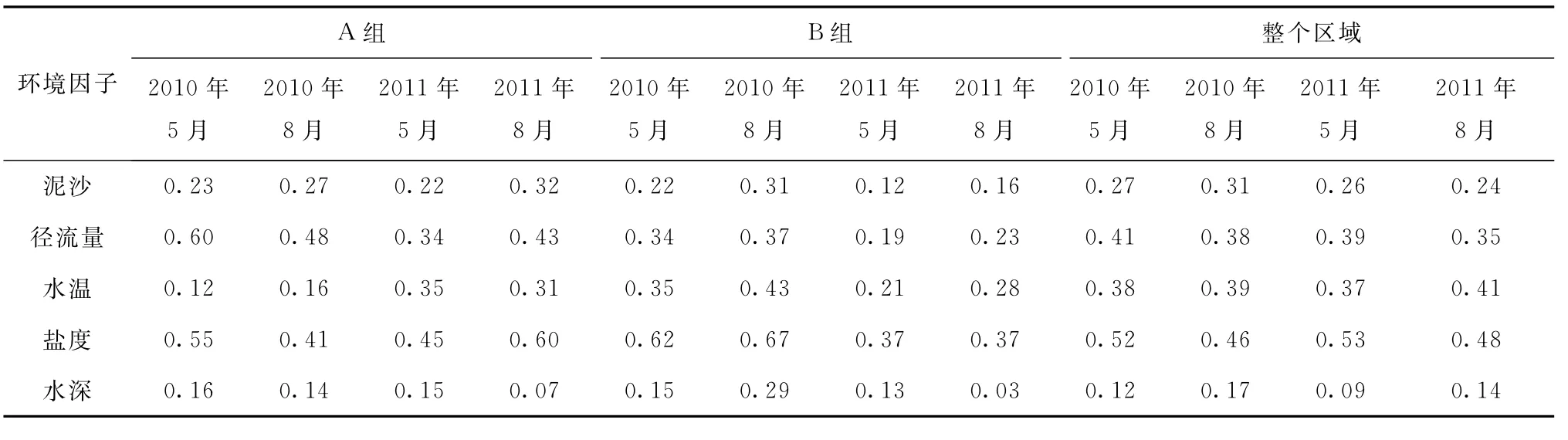
表5 不同時(shí)間群落的總豐度與環(huán)境因子的spearman系數(shù)(r)
4 討論
4.1 種類組成及多樣性
河口由于特殊的地理?xiàng)l件,受徑流和潮流的交匯作用,使得魚類浮游生物群落普遍的特征是擁有少量高豐度的優(yōu)勢(shì)種和眾多的稀有種[15]。而長江口各種魚類的豐度相差很大,魚類居群由少數(shù)豐度大的魚種和多數(shù)豐度小的魚種構(gòu)成[16]。本次調(diào)查顯示鳀科種類的仔稚魚占據(jù)了整個(gè)區(qū)域絕大部分的水域,尤其是該科屬的鳀、鳳鱭和康氏小公魚等魚種IRI均值大于600,對(duì)總豐度貢獻(xiàn)最大,占總捕獲量的80.8%。顯示出鳀科魚類是長江河口及近岸水域最具代表性的種。這與以往學(xué)者關(guān)于河口性魚類研究相一致[17—18]。
由于種群的自身調(diào)節(jié)作用和原生境條件發(fā)生改變都會(huì)導(dǎo)致物種的演替[19]。而水利工程建設(shè)和污染等人為因素也會(huì)造成河口水域魚類的一部分種類消失,一部分種類新出現(xiàn)的現(xiàn)象[20]。從以往歷史資料來看[21],1999年和2001年5月本水域采集到8 808尾仔稚魚,32個(gè)種類,IRI值大于100的種類包括:鳳鱭、鳀、松江鱸Trachidermus fasciatus、白氏銀漢魚Atherina blee犽eri和六絲矛尾鰕虎魚Chaeturichthys hexanema,而本研究中的仔稚魚種類組成與前者有一定差異,數(shù)量略少,尤其是作為優(yōu)勢(shì)種類的松江鱸魚和白氏銀漢魚,已經(jīng)完全消失[22]。這可能與前者調(diào)查時(shí)間正處于長江三峽大壩建設(shè)期,對(duì)于長江下游河口水域而言,其徑流量和泥沙量的調(diào)控能力還未完全顯現(xiàn)。而本次調(diào)查時(shí)間則是三峽大壩已經(jīng)實(shí)現(xiàn)了3次高水位蓄水工程,長江整個(gè)徑流量的調(diào)控能力增加,特別是泥沙輸入量大幅度減少[23]。三峽水庫蓄水后,影響徑流分布,從而改變了長江口及其鄰近海域的理化和生物因子的原因,由于河口環(huán)境條件波動(dòng)較大,魚類浮游生物的種類、數(shù)量、優(yōu)勢(shì)種的結(jié)構(gòu)也會(huì)相應(yīng)地發(fā)生變動(dòng),并且部分魚類的生殖時(shí)間上也出現(xiàn)了相應(yīng)的改變[16]。
物種多樣性指數(shù)反映了種類數(shù)和種間個(gè)體數(shù)量的均衡性[4]。本調(diào)查Shannon-Weaver指數(shù)的差異性表明了群落間物種之間的不均衡性,而這種不均衡性主要是由鳀和康氏小公魚2個(gè)優(yōu)勢(shì)種在豐度上的減少和出現(xiàn)頻率下降引起的(見表1)。物種多樣性過低與增加的環(huán)境擾動(dòng)和不斷變化的非生物環(huán)境條件有一定關(guān)系[1]。根據(jù)物種多樣性指數(shù)評(píng)價(jià)水質(zhì)狀況的標(biāo)準(zhǔn),當(dāng)指數(shù)值為0~1之間,表示水質(zhì)受到重污染[24]。本次多樣性指數(shù)在0.17~0.41之間,表明該區(qū)域水質(zhì)狀況不容樂觀。長江下游水域沿岸多種工業(yè)污染源的排放日益加重,嚴(yán)重污染著河口及鄰近水域的水質(zhì)環(huán)境,同時(shí)過度捕撈作業(yè),對(duì)于物種的生存也造成了一定的影響。
4.2 群落結(jié)構(gòu)環(huán)境影響因素
河口的環(huán)境具有海洋和淡水兩種特性,生物生態(tài)習(xí)性多樣。經(jīng)聚類分析,本調(diào)查水域仔稚魚群落結(jié)構(gòu)可劃分為2個(gè)站位組,分別是低鹽度,高濁度水域的長江河口淡水區(qū)和高鹽度、深水區(qū)的長江口外咸水區(qū)。淡水區(qū)多集中于淡水和半咸水性種類,咸水區(qū)則多為近岸和沿海性種類。可見,在環(huán)境因子中,水溫、鹽度及水系與不同魚類種群有著重要的相互聯(lián)系[25]。
大多數(shù)研究表明,鹽度是決定河口區(qū)魚類空間分布的主導(dǎo)因子之一[26]。本研究發(fā)現(xiàn),低鹽度控制的長江口門內(nèi)(1~16號(hào)站)水域和外海水控制的口門外水域(17~30號(hào)站)的群落結(jié)構(gòu)存在較明顯的差異。長江口內(nèi)常見種以淡水魚類和咸淡水魚類(主要為鯉科和鳀科)為主;口門外水域主要以沿岸性和海洋性種(主要為石首魚科和鰕虎魚科)為主;這可能是由于所處水域鹽度條件的不同所造成。本研究也發(fā)現(xiàn),上述兩大群落的總豐度與鹽度的相關(guān)性較高。而在河口生態(tài)系統(tǒng)中,鹽度區(qū)域性變化十分顯著[27]。
魚類浮游生物群落結(jié)構(gòu)與環(huán)境因子有著密切的關(guān)系。河口區(qū)的魚類浮游生物群落多樣性季節(jié)性變動(dòng)一般都大于年際間的變化[28],本調(diào)查各航次群落相似性指數(shù)表明了同一年季節(jié)性相似性較小(0.38 ~0.40),而年間群落結(jié)構(gòu)相似性較大,相似性指數(shù)在0.50~0.55之間,符合上述河口群落多樣性的季節(jié)變動(dòng)特征。河口魚類浮游生物群落多樣性的季節(jié)變動(dòng)同產(chǎn)卵區(qū)水體環(huán)境的季節(jié)變化對(duì)魚類產(chǎn)卵活動(dòng)的影響有一定的關(guān)系[29]。由于三峽蓄水后,庫區(qū)季節(jié)性調(diào)水至長江下泄量減少,泥沙大部分都儲(chǔ)存在庫內(nèi),河口沖淡水面積減小,減弱了陸源營養(yǎng)物質(zhì)的輸送,海水的自凈能力降低,進(jìn)而影響了河口仔稚魚的分布和群落結(jié)構(gòu)[16]。本研究中也發(fā)現(xiàn)兩大群落組總豐度與泥沙、鹽度和徑流等環(huán)境因子的相關(guān)性分析均顯示出鹽度季節(jié)性差異顯著的特點(diǎn),體現(xiàn)了河口區(qū)域環(huán)境因子季節(jié)性變化的特征(見表5)。
隨著現(xiàn)代工農(nóng)業(yè)的發(fā)展,長江水域富營養(yǎng)化日益嚴(yán)重,并且由于三峽庫區(qū)的調(diào)水,長江徑流減少,污染物逐步向近岸邊緣推移,長江口的流場、溫鹽場和峰面的改變,都將導(dǎo)致河口魚類的棲息環(huán)境的改變,對(duì)于魚類群體的補(bǔ)充資源量終將造成一定的影響[30]。
致謝:本次生物標(biāo)本的采集得到了東海水產(chǎn)研究所晁敏博士、史赟榮博士以及上海海洋大學(xué)碩士畢業(yè)生李聰、黃厚見、廖勇、吳慶元和牛俊翔同學(xué)的大力支持和幫助!
[1] Barletta M,Barletta-Bergan A,Saint-Paul U,et al.The role of salinity in structuring the fish assemblages in a tropical estuary[J].Journal of Fish Biology,2005,66:45-72.
[2] Barletta-Bergan A,Barletta M,Saint-Paul U.Structure and Seasonal Dynamics of Larval Fish in the CaetéRiver Estuary in North Brazil [J].Estuarine,Coastal and Shelf Science,2002,54:193-206.
[3] 羅秉征,沈煥庭,等.三峽工程與河口生態(tài)環(huán)境[M].北京:科學(xué)出版社,1994.
[4] 朱鑫華,劉棟,沙學(xué)坤.長江口春季魚類浮游生物群落結(jié)構(gòu)與環(huán)境因子的關(guān)系[J].海洋科學(xué)集刊,2002,5(44):169-179.
[5] 劉淑德,線薇微.長江口及其鄰近水域魚類浮游生物群落的時(shí)空格局[J].生物多樣性,2009,17(2):151-159.
[6] 蔣玫,沈新強(qiáng),陳蓮芳.長江口及其鄰近水域魚卵、仔魚的種項(xiàng)組成與分布特征的研究[J].海洋學(xué)報(bào),2006,28(2):171-174.
[7] Elliott M,HemingwayKL.Fishes in Estuaries[M].London:Wiley-Blackwell,2002.
[8] 趙傳薩,張仁齋.中國近海魚卵與仔魚[M].上海:上海科學(xué)技術(shù)出版社,1985.
[9] 中國水產(chǎn)科學(xué)研究院東海水產(chǎn)研究所,上海市水產(chǎn)研究所.上海魚類志[M].上海:上海科學(xué)技術(shù)出版社,2001.
[10] 中華人民共和國水利部.2010年中國河流泥沙公報(bào)[M].北京:中國水利水電出版社,2011.
[11] 中華人民共和國水利部.2011年中國河流泥沙公報(bào)[M].北京:中國水利水電出版社,2012.
[12] 趙志模,周新遠(yuǎn).生態(tài)學(xué)引論[M].重慶:科學(xué)技術(shù)文獻(xiàn)出版社重慶分社,1984.
[13] 鄧景耀,金顯仕.萊州灣及黃河口水域漁業(yè)生物多樣性及其保護(hù)研究[J].動(dòng)物學(xué)研究,2002,21(2):76-82.
[14] ClarkKR,Warwick R M.Changes in marine communities:An approach to statistical analysis and interpretation[M].Plymouth:Primer-E Ltd,2001.
[15] Berasategui A D,Acha E M,F(xiàn)erna′ndez Araoz N C.Spatial patterns of ichthyoplankton assemblages in the R?'o de la Plata Estuary(Argentina-Ur uguay)[J].Estuarine,Coastal and Shelf Science 2004,60:599-610.
[16] 單秀娟,線薇薇,武云飛.三峽工程蓄水前后秋季長江口魚類浮游生物群落結(jié)構(gòu)的動(dòng)態(tài)變化初探[J].中國海洋大學(xué)學(xué)報(bào),2005,35(6):936-940.
[17] Jiang M,Shen X Q,Li L,et al.Annual variability of ichthyoplankton in the Yangtze River estuary of China from August 2002 to 2009 [J].Oceanological and Hydrobiological Studies,2013,42:59-69.
[18] Macho G,Vázquez E,Giráldez R,et al.Spatial and temporal distribution of barnacle larvae in the partially mixed estuary of the Ría de Arousa(Spain)[J].Journal of Experimental Marine Biology and Ecology,2010,392:129-234.
[19] Wiebke J B,Janet T D A.Ichthyoplankton dynamics and biodiversity in the Gulf of Alaska:Responses to environmental change[J].Ec-ological indicators 2008,8:292-302.
[20] Autumn O,Scott N.Increasing nutrient concentrations and the rise and fall of a coastal fishery:a review of data from the Nile Delta,E-gypt[J].Estuarine,Coastal and Shelf Science,2008,77(3):308-319.
[21] 劉淑德,線薇微,劉棟.春季長江口及其鄰近海域魚類浮游生物群落特征[J].應(yīng)用生態(tài)學(xué)報(bào),2008,19(10):2284-2292.
[22] 張衡,朱國平,陸健健.長江河口濕地魚類的種類組成及多樣性分析[J].生物多樣性,2009,17(1):76-81.
[23] 陳勇,韓震,楊麗君,等.長江口水體表層懸浮泥沙時(shí)空分布對(duì)環(huán)境演變的響應(yīng)[J].海洋學(xué)報(bào),2011,34(1):145-152.
[24] 張景平,王小平,江志堅(jiān),等.珠江口海域污染的水質(zhì)綜合污染指數(shù)和生物多樣性指數(shù)評(píng)價(jià)[J].熱帶海洋學(xué)報(bào),2010,29(1):69-76.
[25] 陳永俊,林龍山,廖運(yùn)志,等.白令海和楚科奇海魚類種類組成及其對(duì)生態(tài)環(huán)境變化的響應(yīng)[J].海洋學(xué)報(bào),2013,35(2):113-125.
[26] Krumme U.Patterns in tidal migration of fish in a Bra-zilian mangrove channel as revealed by a split-beam echo-sounder[J].Fisheries Research,2004,70:1-15.
[27] Pombo L,Elliott M,Rebelo J E.Changes in the fish faunal of the Ria de Aveiro estuarine lagoon(Portugal)during the twentieth century[J].Journal of Fish Biology,2002,61:167-181.
[28] Blaber S J M,Cyrus D P,Albaret JJ,et al.Effects of fishing on the structure and functioning of estuarine and near shore ecosystems[J]. Journal of Marine Science,2000,57:590-602.
[29] Hernández-Miranda E,Palma A T,Ojeda F P.Larval fish assemblages in nearshore coastal waters off central Chile:temporal and spatial patterns[J].Estuarine,Coastal and Shelf Science,2003,56:1075-1092.
[30] 張怡輝.三峽工程對(duì)長江口水動(dòng)力及污水輸運(yùn)的影響[D].大連:大連理工大學(xué),2011.
Structure of community and biodiversity of larvae fish in spring and autumn 2010—2011 in Yangtze River Estuary and its adjacent waters
Jiang Mei1,Li Lei1,Shen Xinqiang1,Quan Weimin1
(1.East China Sea Fisheries ResearchInstitute,Chinese Academy of Fishery Sciences,Shanghai 200090)
Univariate and multivariate statistics were used to analyze the structure of larvae fish community based on the data collected from the surveys in Yangtze River Estuary and its adjacent waters in spring and autumn 2010—2011.A total of 27 species belonging to 5 orders,14 families were captured from four surveys.The dominant species mainly contained Engraulis japonicus,Coilia mystus,Stolephorus commersonii,Liza haematocheila,Nibea albiflora,Acanthogobius flavimanus and Pseudolaubuca engraulis.The Shannon-Weaver(H′)value fluctuated greatly in an investigation cycle,which was appreciably lower in 2011 than that in 2010.The year variations of community structure and diversity index of larvae fish were more significant in spring than those in summer.The larvae fish community was clustered into two groups which coresponded with the freshwater area and brackish water area of Yangtze River Estuary,respectively.The abundance of larvae fish in both groups was mainly affected by salinity,indicating that salinity was the major cause of the spatiotemporal difference of larvae fish community in the Yangtze River Estuary.
Yangtze River Estuary;larvae fish;community structure;cluster analysis
P734.22
A
0253-4193(2014)06-0131-07
2013-07-22;
2013-11-02。
國家973項(xiàng)目(G2010CB429005)。
蔣玫(1973—),女,江蘇省鎮(zhèn)江市人,研究員,主要從事早期漁業(yè)生態(tài)研究和海洋環(huán)境影響評(píng)價(jià)工作。E-mail:jiangrose73@163.com
蔣玫,李磊,沈新強(qiáng),等.2010—2011年長江河口及鄰近水域春夏季仔稚魚群落結(jié)構(gòu)及其多樣性的研究[J].海洋學(xué)報(bào),2014,36 (6):131—137,
10.3969/j.issn.0253-4193.2014.06.016
Jiang Mei,Li Lei,Shen Xinqiang,et al.Structure of community and biodiversity of larvae fish in spring and autumn 2010—2011 in Yangtze River Estuary and its adjacent waters[J].Acta Oceanologica Sinica(in Chinese),2014,36(6):131—137,doi:10.3969/j.issn. 0253-4193.2014.06.016
猜你喜歡
我愛學(xué)·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時(shí)刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛(wèi)生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
