小黃魚Larimichthys polyactis體長-體重關系冪指數與產卵群體空間分布相關性研究
2014-06-01 12:30:10劉勇程家驊
海洋學報 2014年6期
關鍵詞:研究
劉勇,程家驊
(1.中國水產科學研究院東海水產研究所農業部東海與遠洋漁業資源開發利用重點實驗室,上海 200090)
小黃魚Larimichthys polyactis體長-體重關系冪指數與產卵群體空間分布相關性研究
劉勇1,程家驊1
(1.中國水產科學研究院東海水產研究所農業部東海與遠洋漁業資源開發利用重點實驗室,上海 200090)
魚類產卵群體的相關研究一直是漁業領域關注的熱點,對魚類的產卵期、產卵場的研究是魚類生態習性研究方向的重要組成部分。本文在前期研究證實魚類體長-體重關系冪指數可以指示魚類產卵期的基礎上,利用2004年4個季節的東海大面積調查數據,嘗試利用同一時間點上的各個調查點冪指數信息來分析魚類產卵場分布特征,結果發現,成熟產卵個體僅在春季出現,而未成熟產卵個體4季均有分布,以此推測小黃魚存在“跳過產卵”現象。另外發現,單靠冪指數大小,不能區分各個產卵成熟階段,也不能區分產卵個體與非產卵個體;而懷卵個體出現站點的冪指數平均值偏低于勻速生長,空間上也具有類似特征,即冪指數相對低的地方一般對應懷卵個體相對集中的水域。本文依據冪指數分布,推斷小黃魚成熟產卵群體有3塊相對集中水域,即舟山漁場近海、濟州島西南側和江外與舟外漁場臨近水域。
體長;體重;冪指數;小黃魚;產卵群體;空間分布
1 引言
魚類產卵群體對資源的補充和可持續發展起到至關重要的作用[1—2]。產卵群體研究內容較為廣泛,包括產卵時期、產卵地點和產卵習性等方面內容,這對把握漁汛、保護補充群體和人工繁殖等方面具有重要指導意義[3—8]。產卵群體的研究離不開調查取樣,但此過程涉及范圍大且工作繁重。一般來說,首先是開展大面積調查采樣,然后進行室內樣品分析;對獲取樣品生物學特征逐個分析,其中關鍵的一步是性腺成熟度的觀察與定級,此步不僅需要解剖魚體、尋找性腺體,還需根據經驗判斷定級[9],因此工作繁瑣而且涉及的主觀因素較多,結果不穩定。相比較來說,魚的體長和體重測定的過程結果較為客觀,且在前期研究[10]中發現,體長-體重關系中的冪指數對產卵期有較好的指示作用。本文以2004年小黃魚4個季度的調查數據為例在該方面做進一步探討,分析冪指數是否可以用來判斷產卵群體的空間分布特征。

表1 采樣數據量統計
2 材料和方法
2.1 材料來源
小黃魚生物學測定資料來自2004年東海區漁業資源4次大面積定點調查,共122個站點、3 666尾小黃魚的體長和體重測定數據,各季數據量詳見表1。調查船為雙拖漁輪,功率為183.25 kW/艘,網具為100目× 4 m,網囊網目為2.5 cm,平均拖速為2 n mile/h。調查范圍為27°00′~34°00′N,122°00′~127°00′E,經、緯度每隔30′設1個站位,格狀均勻設置站位。
2.2 方法
2.2.1 體長-體重關系模型
體長-體重關系為冪函數,如下式:
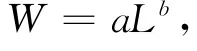
式中,W和L分別為體重(g)和體長(mm);a、b為模型參數。
在對數據進行分析時,剔除偏離整體趨勢的個別值。根據前述函數關系,對各個批次的體長、體重數據進行線性回歸擬合,計算冪指數b值。
2.2.2 等值面插值方法
由于本文是探討通過冪指數b值的分布情況分析推斷小黃魚的產卵范圍,因此采用了普通克里格(OrdinaryKriging,OK)插值方法,該方法建立在半變異函數理論分析基礎上,是對有限區域內變量進行無偏最優估計的一種插值方法[11]。有研究指出,該方法對局部趨勢模擬方面表現較好[12]。
2.2.3 產卵群體劃分方法
參考前期產卵群體研究方法[13],性腺發育處于初期的Ⅰ~Ⅱ期個體予以忽略;把性腺完全成熟、即將或正在產卵的Ⅴ期個體記為B期成熟個體;把早于Ⅴ期、包括Ⅲ期到Ⅳ期的個體記為A期未成熟個體;把Ⅴ期以后、包括產卵后的Ⅵ期和吸收恢復階段(成熟度屬于Ⅱ期)個體[14]記為C期產完卵個體。
2.3 使用軟件
數據處理采用R統計軟件[15];OK插值方法是利用Kriging工具包來實現。
3 結果
3.1 四季度不同成熟度產卵群體分布與體長-體重關系冪指數分布之間的關系
本文所劃分的3種不同產卵群體分布情況如圖1~3所示。不同顏色的等值面對應著不同冪指數值;不同大小圓圈代表著不同數量的產卵群體。可以看到A期未成熟個體在四季中均有分布,且在春季出現站位數最多;B期成熟個體僅在春季調查中出現;C期產完卵個體僅在春季和冬季出現。產卵群體空間分布研究的主要目標是尋找產卵場,A期和C期群體的分布情況僅能代表產卵前期和產卵后期的分布狀況,與產卵場直接聯系的群體則是B期的成熟個體,因此研究重點是B期成熟個體的分布特征。
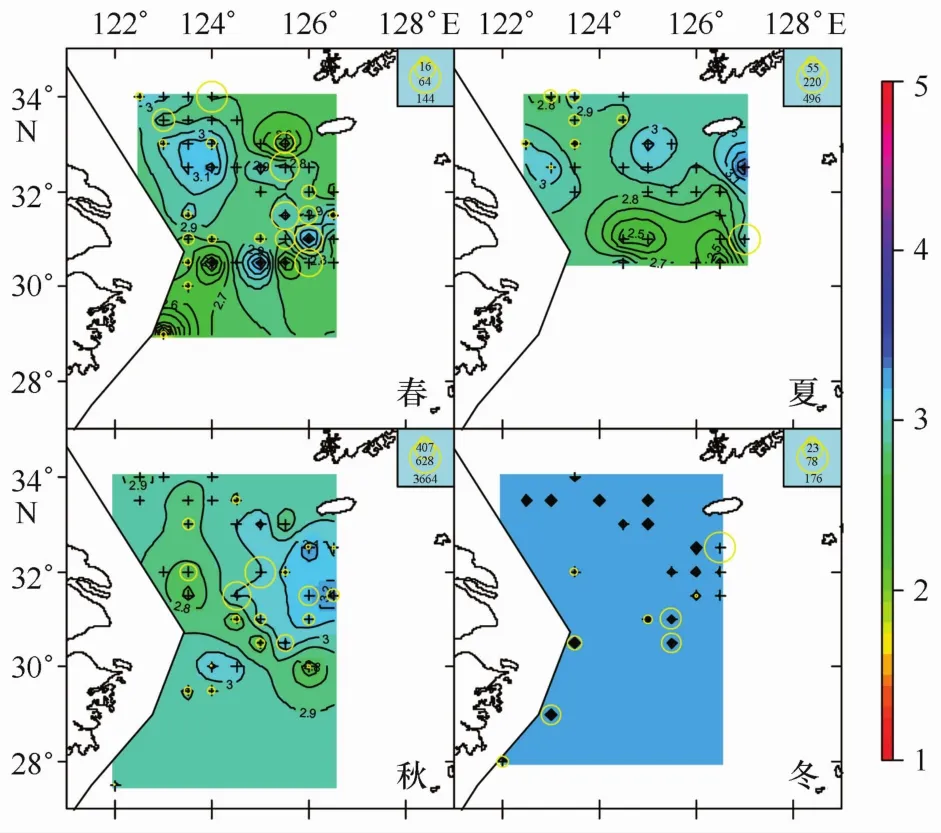
圖1 A期產卵群體四季分布情況
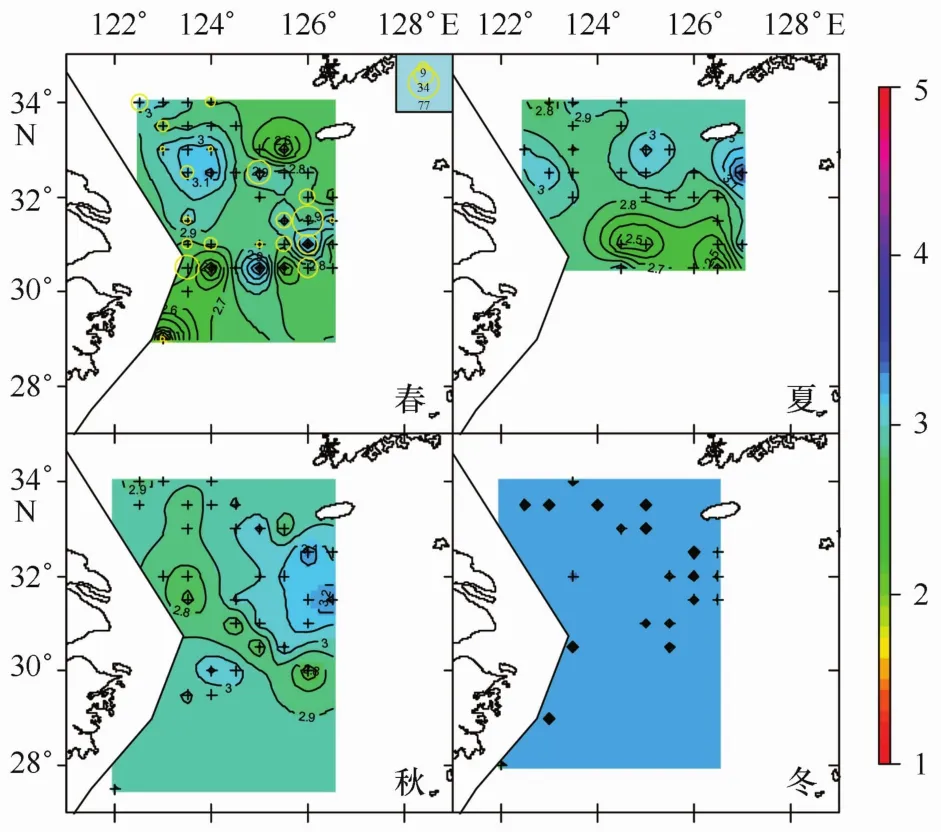
圖2 B期產卵群體四季分布情況
3.2 不同成熟度產卵群體對應的冪指數分布特征
圖4分別對3種產卵群體出現站點及所有調查站點的冪指數分布情況進行比較,并標出分布的主要區間值,上限及下限均是按照10%比例來確定分位數,區間內代表的是80%冪指數的分布情況,可以代表各群體的主要分布特征。由圖4發現,A期未成熟個體所在站點的冪指數分布情況與所有調查站點的分布情況幾乎一致,可見A期個體很難根據冪指數從整體中區分出來。而B期成熟個體就有較明顯地差別,體現在主要區間范圍有一定程度的縮小,范圍為2.77~3.16。C期產完卵個體對應的冪指數有著顯著特征,即冪指數均低于2.89,顯著低于代表勻速生長的冪指數3。
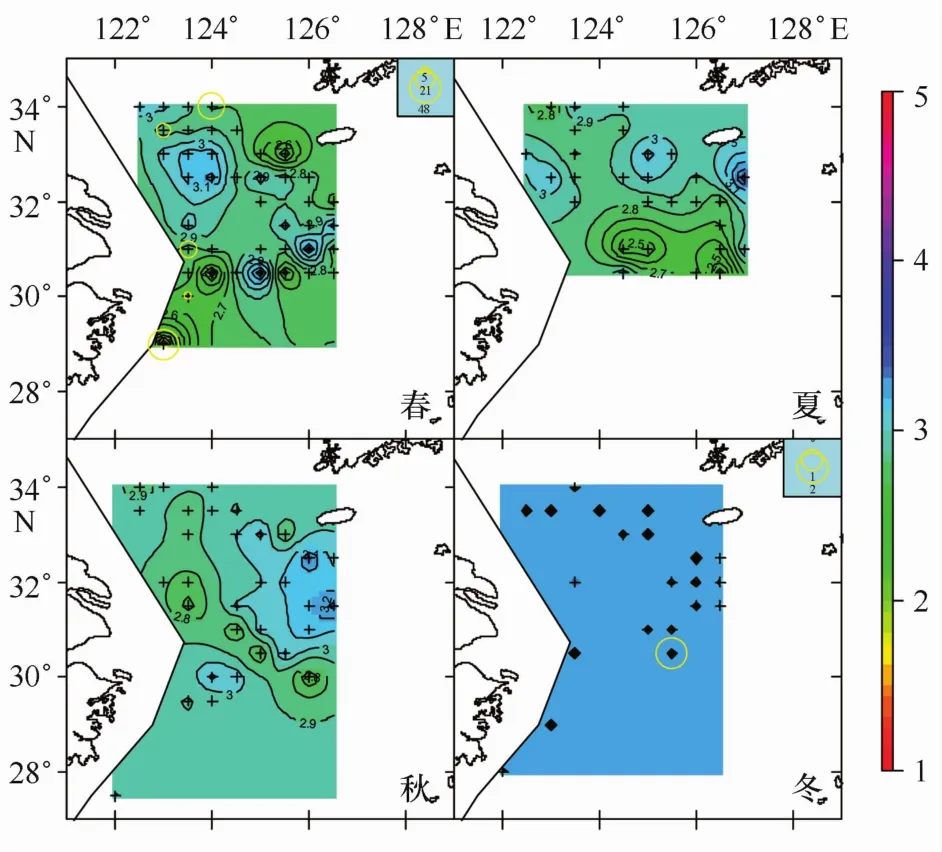
圖3 C期產卵群體四季分布情況
3.3 成熟產卵群體(B期)冪指數代表性分布范圍
根據3.2節分析,B期產卵群體冪指數分布范圍與A期群體或全體相比,差別并不明顯。而觀察B期產卵群體的分布情況(見圖2,春季),可以發現成熟產卵群體出現3個比較明顯的聚集區域,一個是右邊偏下側區域,一個是右上角區域,還有一個是左下角區域,并且這3個區域都對應一個冪指數值相對較低的區域。雖然成熟產卵群體冪指數分布范圍相對廣泛,而大部分成熟產卵個體是否存在一個相對較狹窄的冪指數分布范圍?沿此思路,就不同冪指數范圍內,對應出現站位數及產卵個體數占整體調查的比例進行了統計,結果見表1。比較發現,冪指數b值小于2.9的范圍可以涵蓋六成以上的產卵個體,可以涵蓋半數以上的出現站點,因此本研究以2.9為閾值來分析多數產卵個體的分布范圍。在圖2(春)基礎上,去除2.9以上的區域,剩余的區域應為本文結果所得的產卵區域(圖5)。
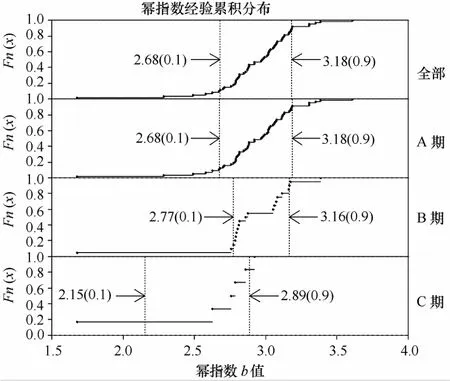
圖4 不同產卵群體的冪指數分布情況
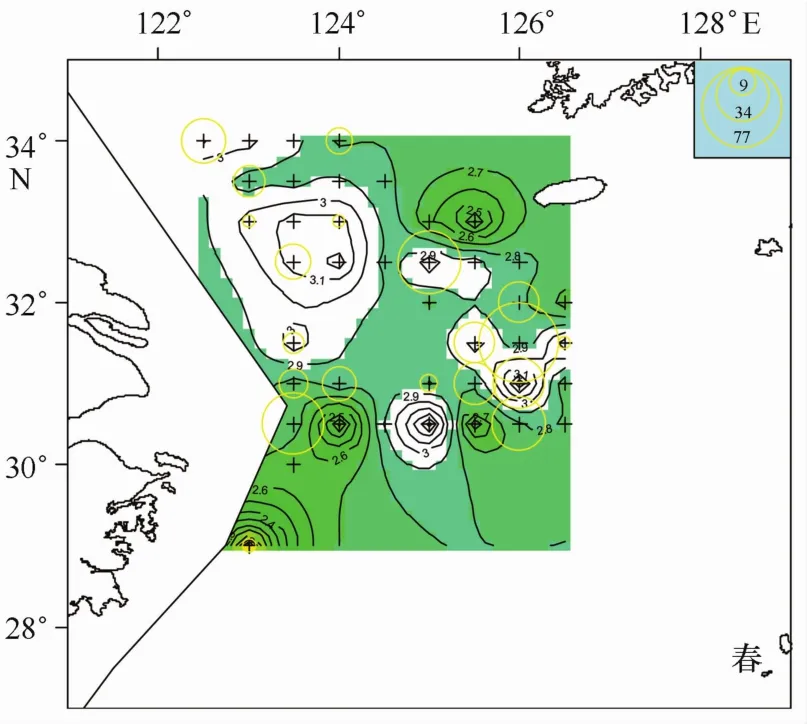
圖5 小黃魚產卵區域推斷(b<2.9)
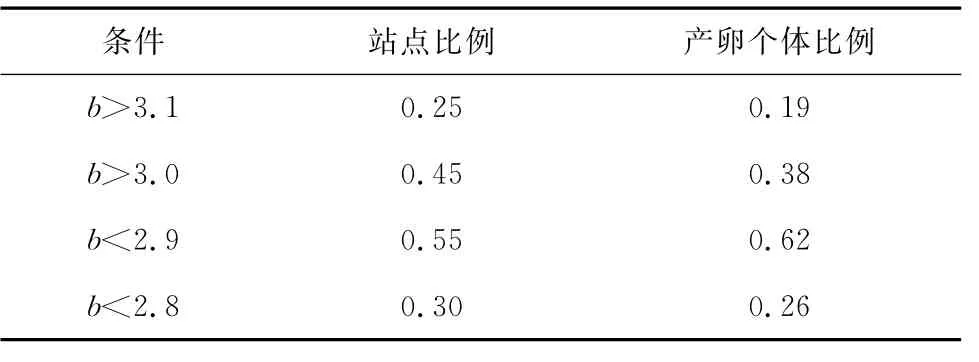
表2 成熟產卵群體不同冪指數范圍的統計數據
4 討論
4.1 不同成熟階段產卵群體四季分布的差別
本研究結果3.1節中,產卵個體僅在4月調查中出現,這與先前研究結果相吻合,即小黃魚的產卵時間主要集中在4-5月份[10,14,16—18]。產過卵個體,在春季和冬季調查中出現;春季出現產過卵個體說明調查時間是小黃魚開始產卵后開始,部分個體已經完成產卵;調查結果中,冬季也出現產過卵個體,冬季應該是小黃魚索餌洄游、開始新一輪產卵周期的季節[3],正常情況下不會出現產過卵個體;經核實僅有一個站點,且僅有一尾魚被判斷為產過卵,這可能是因為初期發育性腺與產完卵性腺外觀近似、導致的產卵期誤判。
而未成熟個體,4個季節均有分布,春季出現的站點和數量均最多。4至5月份是小黃魚的主要產卵期,即使過了產卵期,仍然存在許多未成熟產卵個體,并且在后續的3個季節中均保持一定數量;另外據產卵B期個體僅在春季出現,未在其他季節發現產卵個體,進而可以推斷這些未成熟個體是錯過了產卵期、并未能在當年產卵。這一現象被稱作“跳過產卵(skip spawning)”,即產卵個體因為當年產卵期的水溫不適宜,餌料貧乏,或魚群密度不正常、性別比例失衡,或環境污染等因素影響,從而導致自主放棄當年產卵的行為[19—20]。本研究的結果可以為小黃魚存在“跳過產卵”現象提供一個證據。
4.2 產卵群體與低冪指數區域相關趨勢
冪指數b可以用來判斷魚類是否處于勻速生長[2,21]。魚類不同生長時期因生長發育的不均勻性,即體重和體長增長不均勻,從而帶來冪指數b值的上下波動。據此特點,基于小黃魚年度冪指數b值序列,識別其產卵期的設想已在前期研究[10]中得到了驗證;本研究繼續嘗試基于一個時間點上的冪指數b值空間分布,判別小黃魚產卵群體的分布特征。
根據冪指數b值來推斷小黃魚產卵群體的分布范圍,理想狀態是存在性腺不同成熟階段個體的站點對應的冪指數可以明顯區分,但數據分析的結果并不理想(見圖4),不僅懷卵個體(包括A、B和C期)與全體之間區分不開,而且各性腺成熟階段個體之間區別也不明顯,例如A期對應冪指數范圍包含了B期范圍,C期雖與A、B期相比明顯有偏小的特征,但與兩者之間也存在交叉范圍,不能完全分隔。這也許是因為3種成熟階段的懷卵個體并不是完全分離,例如它們可能同時在一個站點中出現、從而共同對應一個冪指數,因而單純依據冪指數是不能區分各個產卵成熟階段;懷卵個體與全體區分不開,因此也不能依據冪指數區分產卵個體與非產卵個體。
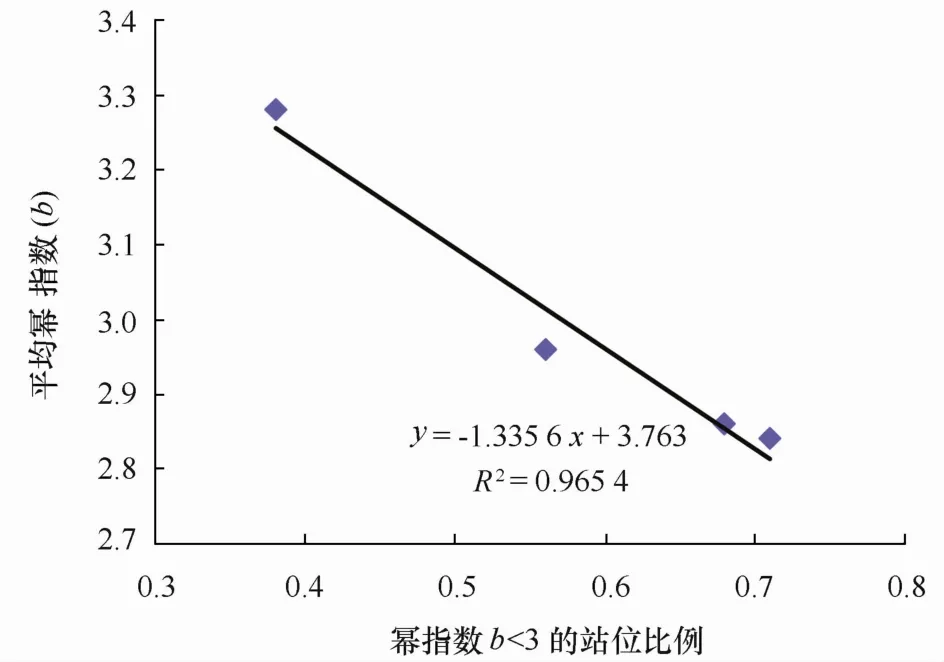
圖6 產卵出現比例與冪指數平均值關系
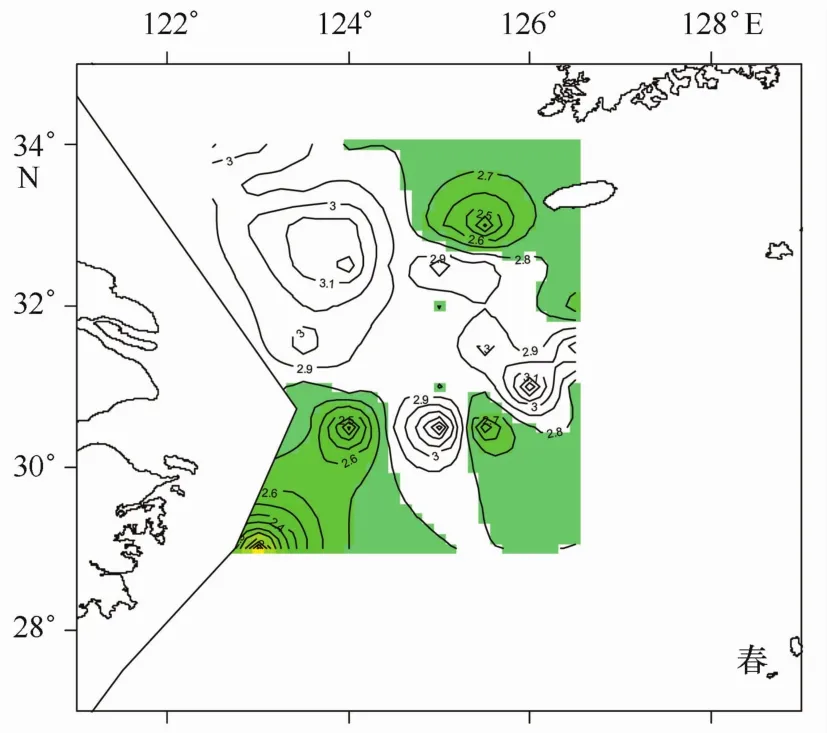
圖7 小黃魚產卵區域推斷(b<2.8)
圖4是把全年數據匯合在一起進行分析,各個季節不同成熟階段個體出現站點組成差別較大,如四季懷卵個體出現站點數分別為23、8、20和9(見表3),各期成熟個體出現的站點數更是差別明顯。為了分離季節、性腺成熟階段因素的相互影響,本文對不同季節、不同成熟階段個體對應冪指數的相關統計特征進行了匯總(見表3),對不同性腺成熟期的季節變化特征進行了分析。4個季節的冪指數平均值,春季對應的是小黃魚產卵季節[4],因而相對較低;夏季的冪指數平均值達到一個低谷,隨后的秋季和冬季逐步上升,冬季的冪指數平均值達到一個較高值,這可能是由于小黃魚在結束產卵后就開始索餌洄游,繼而是越冬洄游,從而其體型發生變化而導致,該結果與前期研究較為一致[4]。觀察表3發現,4個季節的全部冪指數平均值與冪指數值小于3的站點比例呈反比關系,把兩者對應的坐標點畫在同一個坐標系中(見圖7)進行觀察,兩者之間存在著顯著線性關系(p<0.02)。冪指數平均值會隨著冪指數值小于3的站點比例的增多而變小,而這一規律在“懷卵個體”的冬季卻未出現(見表3);相比其它季節,冬季冪指數小于3的站點比例雖然有所增加,冪指數平均值沒降反升;可推斷冬季懷卵個體小于3的冪指數值與勻速生長值3的偏差并不大,均處于低于3的較高水平,可能是因為這些錯過產卵季節的繁殖個體停止了產卵進程、加入了索餌和越冬洄游導致。所有季節3個性腺成熟階段個體出現站點的平均冪指數,除A期的冬季略微高于3外,其余均低于代表勻速生長的冪指數3,可見懷卵個體體形生長已經偏離勻速生長,且冪指數是偏向變低的。這種懷卵個體冪指數平均值偏向低于勻速生長的現象在空間上也有體現,觀察圖1-3,可以看到,冪指數相對較低的水域一般均對應著懷卵個體較多的地方。
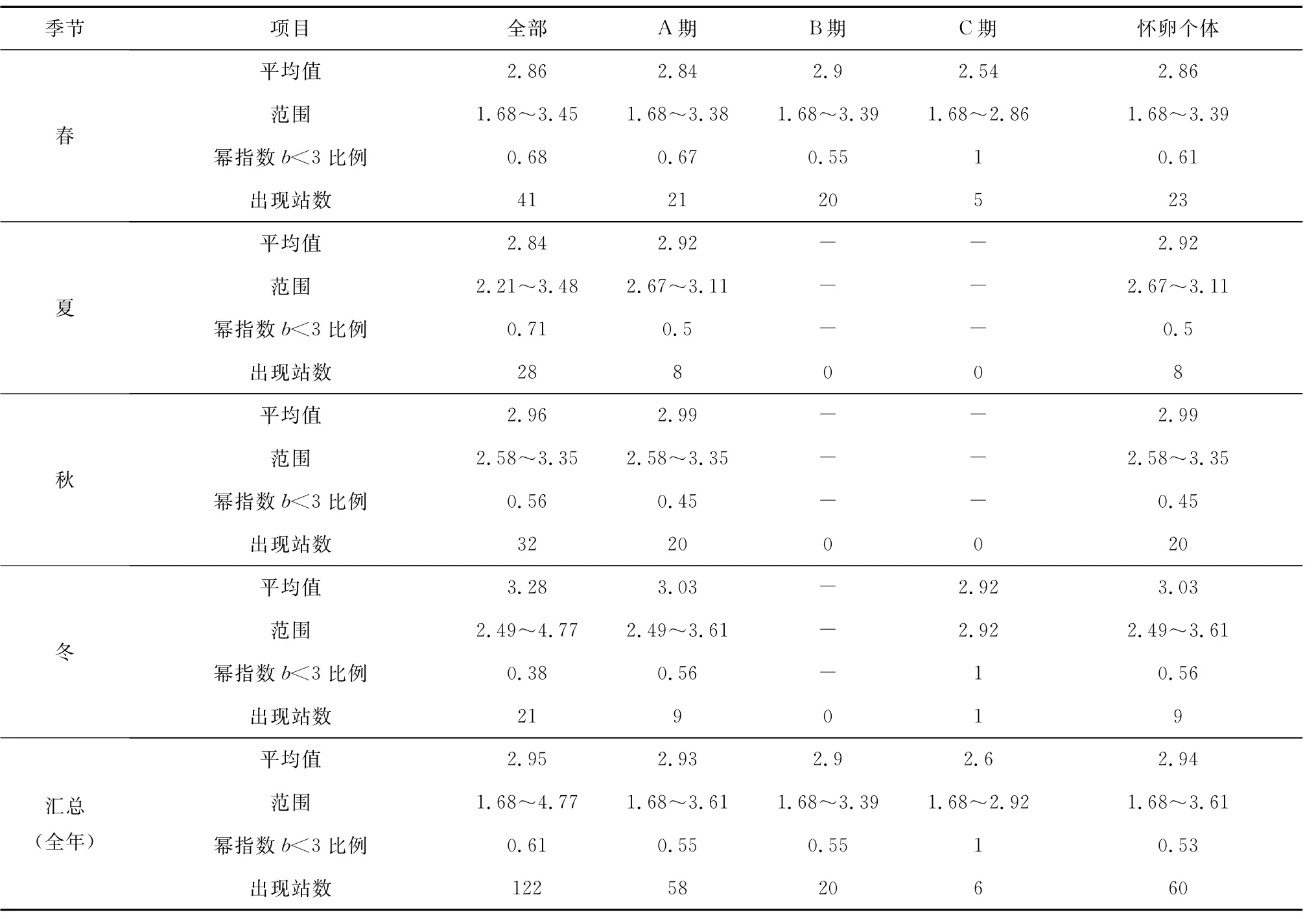
表3 不同季節不同成熟個體冪指數b值特征統計
4.3 3個產卵相對集中水域
根據本文分析結果發現,成熟產卵群體出現水域,或者說是小黃魚產卵水域,調查范圍內相對集中在3塊,一是處于舟山漁場近海水域,二是位于濟州島西南側水域,另一塊是位于江外漁場與舟外漁場臨近部分水域(見圖5)。與歷史文獻調查結果相比較,位于浙江近海、舟山群島附近海域的產卵場[14,17—18,22],與本文分析得到的舟山漁場水域相吻合。而另外兩塊水域,在上述歷史調查文獻中鮮見報道。濟州島西南側水域在2003年調查[13]中同樣有出現,而2003年分布中心相對偏南;比較2003年與2004年成熟產卵群體的分布特征,2004年外海的這兩塊產卵水域很可能是2003年濟州島西南側產卵場分別向北和向南分離出來的兩塊相對集中的水域。連續兩年調查均在外海水域發現成熟產卵群體,是否證明這一水域已經成為小黃魚相對穩定的產卵場有待確定。另外,本研究調查在呂四漁場范圍內未能發現產卵群體聚集中心,而歷史調查中呂四漁場是一個重要的產卵場[14,17—18,22],2003年調查該水域也有產卵群體聚集,什么原因導致2004年產卵群體聚集中心在該水域消失?是當年特殊的氣候或水團等環境因素影響還是該水域小黃魚群系的親體被過度捕撈的原因?這些問題有待進一步調查。
根據冪指數雖然不能把產卵群體與其他群體完全分隔開來(見圖4),但利用上述觀察到的產卵群體對應偏低的冪指數特征(見3.2節),可以依據冪指數分布推斷產卵群體大概的分布聚集水域,這種方法給判斷產卵區域提供了一種新途徑。圖5是依據b< 2.9推斷的小黃魚產卵水域,涵蓋了大多數成熟產卵群體的分布范圍,但很難看出3個相對集中水域,若把冪指數范圍限制到b<2.8(見圖7),3塊聚集水域就自然顯現出來。雖然這3塊水域不能涵蓋多數成熟產卵群體的分布范圍(見表2),但它把小黃魚成熟產卵群體分3塊相對集中水域分布的特征和趨勢非常明了的展現出來,因而根據冪指數分析小黃魚產卵群體分布情況,更容易發現其分布特征和趨勢。
[1] Hilborn R,Walters CJ.Quantitative fisheries stock assessment:choice,dynamics and uncertainty[J].Reviews in Fish Biology and Fisheries,1992,2(2):177-178.
[2] 詹秉義.漁業資源評估[M].北京:中國農業出版社,1995.
[3] 殷名稱.魚類生態學[M].北京:中國農業出版社,1995.
[4] 徐漢祥,劉子藩,周永東.東海帶魚生殖和補充特征的變動[J].水產學報,2003,27(4):322-327.
[5] 程家驊,林龍山,凌建忠,等.東海區小黃魚伏季休漁效果及其資源合理利用探討[J].中國水產科學,2004,11(6):554-560.
[6] 凌建忠,嚴利平,林龍山,等.東海帶魚繁殖力及其資源的合理利用[J].中國水產科學,2005,12(6):726-730.
[7] 張輝,袁興偉,程家驊.東海區小黃魚繁殖模型優化選擇及其管理應用研究[J].中國水產科學,2010,17(6):1300-1308.
[8] 徐漢祥,劉子藩,周永東,等.伏季休漁條件下東海帶魚的親體和補充量關系[J].漁業現代化,2011,38(1):64-69.
[9] 陳大剛.漁業資源生物學[M].北京:中國農業出版社,1997.
[10] 劉勇,程家驊.基于Bootstrap統計方法探討體長-體重關系中的冪指數對小黃魚產卵期的指示作用[J].漁業科學進展,2011,32(2):1-6.
[11] 侯景儒,胡平昭,黃競先,等.實用地質統計學[M].北京:地質出版社,1998.
[12] 張偉,李純厚,賈曉平,等.底棲生物生物量空間插值方法研究[J].海洋通報,2010,29(3):351-356.
[13] 劉勇,嚴利平,程家驊.2003年東海北部和黃海南部外海小黃魚產卵群體的分布特征及其與水溫、鹽度的關系[J].中國水產科學,2007,14 (7):89-96.
[14] 葉昌臣.小黃魚[M]//海洋漁業生物學.北京:農業出版社,1991:164-200.
[15] R Development Core Team.R:A Language and Environment for Statistical Computing[CP/OL].http://www.R-project.org.
[16] 朱樹屏.黃渤海區小黃魚的洄游及有關環境因素[M]//太平洋西部漁業研究委員會第三次全體會議論文集.北京:科學出版,1960:16-24.
[17] 毛錫林,俞積楚,秦憶芹.小黃魚[M]//東海區漁業資源調查和區劃.上海:華東師范大學出版社,1987:339-356.
[18] 俞連福,程家驊.小黃魚[M]//東海大陸架生物資源與環境.上海:上海科學技術出版社,2003:472-487.
[19] Rideout R M,Rose G A,Burton M P.Skipped spawning in female iteroparous fishes[J].Fish and Fisheries,2005,6(1):50-72.
[20] Rideout R M,Tomkiewicz J.Skipped spawning in fishes:More common than you might think[J].Marine and Coastal Fisheries,2011,3(1):176 -189.
[21] Ricker W E.Computation and interpretation of biological statistics of fish populations[J].Bulletin of the fisheries research board of Canada,1975,191:382.
[22] 徐兆禮,陳佳杰.小黃魚洄游路線分析[J].中國水產科學,2009,16(6):931-940.
Study on the correlation between spatial distributions of the spawning groups and the power b in length-weight relation function of small yellow croaker(Larimichthys polyactis)
Liu Yong1,Cheng Jiahua1
(1.Key Laboratory of East China Sea&Oceanic Fishery Resources Exploitation andUtilization,Ministry of Agriculture,China,East China Sea FisheriesInstitute,Chinese Academy of Fishery Sciences,Shanghai 200090,China)
Studies on fish spawning group are always the focus of fisheries science.Spawning periods and areas are important parts of fish ecological habit studying.Power b in exponent relationship between fish body length and weight as spawning period indicator has been tested and proved with one whole year data in previous paper.Based on the above,this study tries to analysis the spatial character of fish spawning groups by power b at a time point with the data collected from a bottom trawl survey in the 4 seasons of 2004 in the East China Sea.The results show that,ripening small yellow croakers appeared only in spring,however immature individuals appeared in all of 4 seasons,which suggests the presence of“skip spawning”phenomenon.Another finding is that different spawning mature stages can not be distinguished only based on the value of power b,neither the difference between the spawning and non-spawning;while the average power b of sites including spawning individuals appeared lower than that in a constant speed growing,and similar characteristics also occurred in the space,where the relatively low power b generally occurred in the waters with relative high concentration of spawning individuals.Based on power b,three areas with relative high concentration of ripening individuals were inferred,including the offshore of Zhoushan fishing ground,the southwest side of Jeju Island and the adjacent water between Jiangwai and Zhouwai fishing grounds.
body-length;weight;power b;small yellow croaker;spawning groups;spatial distribution
S931.9
A
0253-4193(2014)06-0124-07
2013-06-28;
2013-12-03。
農業部專項東海區海洋捕撈基礎信息動態采集分析(2011—2013);農業部專項中日暫定水域漁業資源調查(2011—2013);農業公益性行業科研專項——東海漁業資源評價和增殖養護技術研究與示范。
劉勇(1977-),男,江蘇省東臺市人,副研究員,博士,從事海洋漁業資源與生態學研究。E-mail:liuy@ecsf.ca.cn
劉勇,程家驊.小黃魚Larimichthys polyactis體長-體重關系冪指數與產卵群體空間分布相關性研究[J].海洋學報,2014,36(6):124—130,
10.3969/j.issn.0253-4193.2014.06.015
Liu Yong,Cheng Jiahua.Study on the correlation between spatial distributions of the spawning groups and the power b in length-weight relation function of small yellow croaker(Larimichthys polyactis)[J].Acta Oceanologica Sinica(in Chinese),2014,36(6):124—130,doi:10.3969/j.issn.0253-4193.2014.06.015
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
