大白菜核不育雄蕊敗育超微結構觀察
2014-01-20 07:56:34高宏秀
浙江農業科學 2014年2期
高宏秀,張 瑩
(江蘇徐州生物工程職業技術學院,江蘇徐州 221006)
大白菜核不育雄蕊敗育超微結構觀察
高宏秀,張 瑩
(江蘇徐州生物工程職業技術學院,江蘇徐州 221006)
以大白菜雄性不育近等基因系為材料,運用TEM技術進行雄蕊發育過程中的絨氈層細胞的超微結構研究。結果顯示,在四分體時期,可育花藥絨氈層細胞開始橫向增大,細胞核清晰可見,核仁染色很深,細胞質內出現大小不一致的囊泡;淀粉類物質聚集成啞鈴形、球形等多種形狀。此時絨氈層細胞細胞膜較薄,細胞之間排列緊密;而同時期的不育花藥絨氈層開始膨大,出現許多小空泡,細胞膜明顯變厚,隨后絨氈層向藥腔伸展,出現多層細胞,有的細胞空泡化嚴重;細胞質內部未見細胞核和細胞器,也未見糖類物質。有的細胞局部空泡化,細胞質內含有少量的含油體和線粒體,由于糖類物質較少,細胞質比較清晰。說明絨氈層的結構異常導致無法提供給小孢子發育所需的營養物質,最終導致雄蕊敗育。
大白菜;核不育;超微結構觀察
雄性不育(malesterility,MS)是指植物在遺傳、環境(溫度和光照等因素)等因子的影響下雄性器官退化,發育不良或花粉、精子敗育不能夠行使生育能力而雌蕊卻發育正常的現象[1]。這種現象在植物界普遍存在。在高等植物中,約有43科162屬320種約620個品種或種間雜交種具有雄性不育現象[2-3],并在新的科學技術手段的運用過程中,創造了一批人工雄性不育體。雄性不育系主要是利用天然具有雜種優勢的植株作為母本,省去人工去雄的煩瑣工作,便于大規模的雜交制種,為生產上大規模利用雜種1代優勢創造有利條件,這樣不但能夠生產出具有較高純度的商業雜交種,還具有較強的知識產權保護功能。自1921年Bateson等首先在亞麻(Linumusitatissimum)中發現雄性不育以來,植物雄性不育機理的研究一直是一個熱門課題。
大白菜(BrassicacampestrisL.ssp.chinensis Makino)原產于我國北方地區,是十字花科蕓薹屬蕓薹種葉用蔬菜,素有“菜中之王”的美稱,其栽培面積和消費量在我國居各類蔬菜之首。大白菜為兩性花異花授粉,雜種優勢十分明顯。由于花器官比較小,單花結籽量很少,而單位面積的播種量又較大,人工授粉制種難以大面積地實現其雜種優勢的商業化應用。大白菜的遺傳背景比較簡單,不育株和可育株僅受1對隱性基因控制,可在較短時間內將顯性目的基因固定到雜交1代上,且F1代的營養器官是大白菜育種利用的主要部位,可不經過育性恢復直接用于生產,因此大白菜雄性不育系利用一直被認為是目前F1代雜種生產的理想途徑。
關于核基因雄性不育系的遺傳機制的早期研究認為,不育性是由核內一對隱性基因控制,也有人提出是由1對顯性基因控制。通過測交篩選法發現,這兩類不育材料都找不到完全的保持系,只能獲得不育株率穩定在50%的兩用系。用兩用系配制商業雜交種,必須在開花前拔除可育株,增大了制種成本;在大面積制種時,雜種純度難以保證。20世紀90年代,大白菜核基因雄性不育研究工作有了重大的突破。100%不育植株率的核不育材料選育成功,并對該類核不育材料的遺傳機制進行摸索,獲得了一系列遺傳假說。
張書芳等[4]通過對小青口、二青幫、青幫河頭等品種的雜交篩選,獲得了不育株率穩定在50%左右的不育系統,但這種兩用系(既可作不育株也可作可育株)在生產應用中面臨著不育株育性不穩定的問題,需進一步改良后才有可能應用于生產。VanDerMeer等[5]在用大白菜品種Granaat和Pavchoi雜交的F1代中發現不育株,用Monument品種回交育成了顯性控制不育性的核不育材料。張書芳等[4]在農家品種萬泉青幫中發現了顯性不育基因(Sp)及其顯性上位基因(Ms),并找到了臨時保持系,育成了具有100%不育株率的雄性不育系88-1A。魏毓棠等[6]也獲得了類似的核不育體系。馮輝[7]等獲得雄蕊退化徹底、不育株率和不育度均為100%的復等位基因控制的大白菜核不育材料,并提出了大白菜核基因雄性不育“復等位基因遺傳假說”。該類不育系既不存在細胞質雄性不育系的胞質負效應,也不存在制種時撥除可育株的問題,是一種較為理想的雄性不育系[8]。核不育花藥的細胞學研究主要就是為了研究花粉敗育的過程,這樣可將核不育分子水平研究和個體調控育性變化的研究聯系起來,有利于全面認識核不育機理。
本研究以大白菜核不育近等基因系為材料,運用電鏡超薄切片技術手段對材料花藥的發育進行研究,預期獲得大白菜核不育花藥的確切敗育時間以及敗育特點,并為大白菜核不育的細胞凋亡檢測提供細胞學依據。
1 材料與方法
1.1 試驗材料
大白菜核不育近等基因系L3,可育株基因型MsfMs,不育株基因型MsMs,遺傳穩定,系統內兄妹交后代育性保持1∶1分離,由云南農業大學大白菜課題組選育而成。
1.2 試驗方法
透射電鏡切片觀察于盛花期,按花蕾在花序上的位置連續取樣,每樣設3個重復,將材料迅速投入2.5%戊二醛固定液中固定。(1)前固定。50mmol·L-1pH值7.4的磷酸鈉緩沖液,含2.5%的戊二醛。室溫固定6h,磷酸鈉緩沖液漂洗3次,每次30min。(2)后固定。1%四氧化鈉,50mmol·L-1pH值7.4的磷酸鈉緩沖液中, 4℃固定12h,磷酸鈉緩沖液漂洗3次,每次30 min。(3)脫水。用30%,50%,70%,80%, 90%的酒精逐級脫水,每級20min,100%酒精脫水3次,每次30min。(4)滲透。花藥放在酒精∶樹脂為2∶1中,室溫下滲透2h;酒精:樹脂為1∶1中滲透5h;酒精∶樹脂為1∶2中滲透12h;純樹脂中滲透48h,中間換1次新鮮配制的樹脂。(5)包埋和聚合。用牙簽輕輕挑起花藥,然后放入預先裝滿包埋樹脂的包埋板上的小凹槽中,并使組織塊位于小凹槽的最右端,按照切片的要求調整組織塊的方向,在恒溫箱中60℃條件下聚合48h。(6)切片。用LeicaEMTRIM修塊機修快,在LeicaEMUC6超薄切片機上切片,厚度90nm,用75目銅網作載網撈取連續切片。(7)染色。醋酸雙氧鈾和檸檬酸鉛在室溫下雙染色各20min。(8)觀察。載網于JEM-1200EXⅡ型透射電鏡下80kV觀察并拍照。
2 結果與分析
2.1 可育花藥發育過程結構觀察
通過前期對大白菜可育花藥的石蠟切片觀察,在四分體之前絨氈層細胞發育正常。進一步用透射電鏡切片觀察大白菜核可育花藥雄蕊四分體時期至花粉成熟期絨氈層的結構。
在四分體時期,隨著藥腔的發育,絨氈層細胞開始橫向增大,細胞核清晰可見,核仁染色很深,細胞質內出現大小不一致的囊泡;淀粉類物質聚集成啞鈴形、球形等多種形狀。此時絨氈層細胞細胞膜較薄,細胞之間排列緊密(圖1中A,B)。在單核小孢子時期,絨氈層細胞已有局部橫向伸長,細胞質清晰,細胞質內有液泡、小囊泡、淀粉粒等細胞器,細胞核染色較淺,核仁染色較深,液泡可見(圖1中C)。隨后絨氈層細胞縱向皺縮,橫向擴張,發展成為細長型細胞,細胞開始出現大量的空泡化現象,細胞內部的糖類物質積累增多,細胞壁已經不明顯,但是還存在明顯的細胞輪廓,同時細胞間隙也不斷加大(圖1中D)。最后在花粉即將成熟時,絨氈層則降解明顯,絨氈層已經沒有明顯細胞輪廓,絨氈層含有豐富的營養物質,細胞內存在淀粉粒和核小體(圖1中E),當花粉完全成熟時,絨氈層完全降解,花藥內壁細胞成為一個空細胞,細胞內部細胞質、細胞器都基本消失,細胞壁也變薄(圖1中F)。
2.2 不育花藥發育過程結構觀察
通過前期對大白菜可育花藥的石蠟切片觀察,不育花藥的絨氈層在四分體時期異常發生,隨后進一步用透射電鏡切片觀察大白菜核敗育花藥四分體時期至花粉成熟期絨氈層的結構。
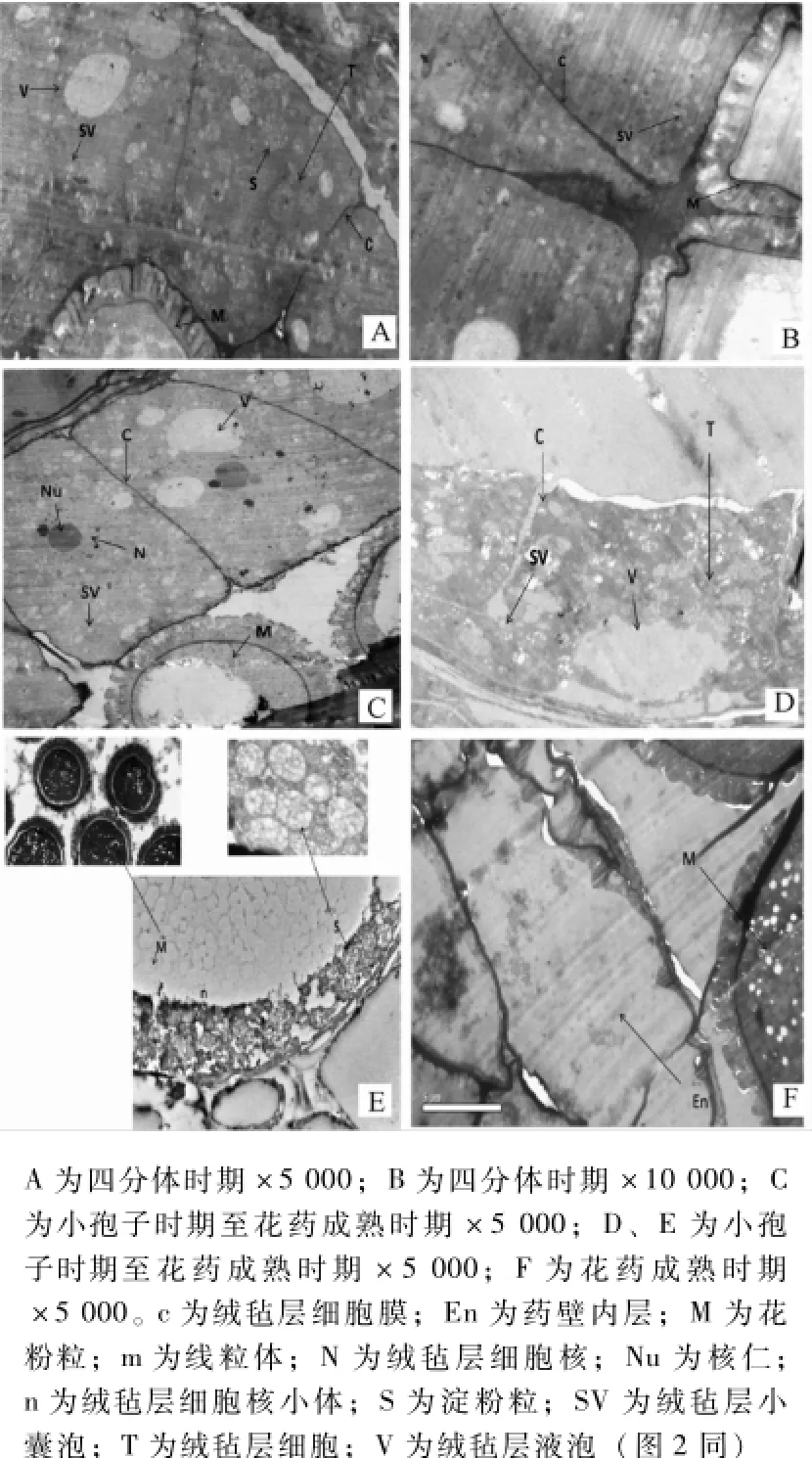
圖1 可育花藥超微結構

圖2 不育花藥超微結構
在四分體時期,藥絨氈層開始膨大,出現許多小空泡,細胞質中可以看見小囊泡、含油體等細胞器,細胞核和核仁可見,但染色較淺,此時細胞壁明顯變厚(圖2中A)。隨后的小孢子向成熟期發育過程中,絨氈層向藥腔擴增,出現多層細胞,有的細胞空泡化嚴重,細胞質內部已經沒有細胞核和細胞器了,也未見糖類物質;而有的細胞局部空泡化,細胞質內含有少量的含油體和線粒體等細胞器,但是糖類物質較少,細胞質比較清晰(圖2中B)。在小孢子發育后期,絨氈層細胞的空泡化更加嚴重,細胞被空泡占據大部分空間,細胞質較少,可見個別線粒體,但其他細胞器和質體已經很難發現了,同時細胞也開始變得不規則,在接下來的發育中絨氈層也開始皺縮,細胞變得很不規則,細胞的細胞壁開始變薄,細胞內部得高度空泡化,基本不具備活細胞的特征(圖2中C)。至花粉成熟期,絨氈層完全皺縮退化,已觀察不到絨氈層細胞了,此時花粉粒也降解完全,花藥全部敗育(圖2中D)。
3 小結與討論
絨氈層處于花藥結構的最內層,與小孢子直接接觸,所以絨氈層細胞對小孢子的形成和發育起到至關重要的作用。對大多數植物而言,絨氈層的功能主要是產生和輸送給小孢子正常發育所必需的酶、生長素和營養物質。從結構水平來講,絨氈層的發育特征表現為順序的細胞結構的消除。絨氈層的分化及之后的退化與減數分裂后的花藥發育成熟相協調一致。從細胞水平來講絨氈層細胞是一種特殊的分泌細胞,絨氈層在小孢子母細胞完成減數分裂后能適時地分泌胼胝質酶,分解四分體周圍的胼胝質壁,促使其釋放小孢子。由于絨氈層合成與分泌胼胝質酶具有時間和空間特異性,在有花植物花粉的正常發育中起著極其重要的決定作用。所以絨氈層的任何變化,都可能影響到小孢子的正常發育,從而導致雄性不育。正常的絨氈層細胞,在減數分裂時期細胞之間連接松弛,減數分裂結束后開始解體,隨著花粉的逐步成熟而逐漸退化,僅留殘片。絨氈層發生功能失常,常被認為是花粉敗育的主要原因。如絨氈層提早成熟或解體、絨氈層增生或肥大、絨氈層不解體或推遲解體均可引起雄性不育。對本試驗材料在電鏡切片中發現,在四分體時期絨氈層開始膨大,出現空泡化,中層細胞開始退化,變成細長中空的細胞,細胞壁明顯變厚。隨后的小孢子向成熟期發育過程中,絨氈層向藥腔擴增,出現多層細胞,有的細胞空泡化嚴重,細胞質內部已經沒有細胞核和細胞器了,也沒有糖類物質;有的細胞局部空泡化,細胞質內含有少量的含油體和線粒體等細胞器,但是糖類物質較少,細胞質比較清晰,這和可育性好的花藥同時期的結構差異較大,最明顯的標志就是供給小孢子發育的糖類物質基本不存在,當然也無法釋放給小孢子,所以絨氈層結構的異常以及其部分功能的喪失,最終引起小孢子敗育。
大白菜核不育雄蕊敗育與花藥絨氈層細胞結構異常密切相關,絨氈層空泡化并異常膨大,細胞內的囊泡、線粒體、糖類物質都不斷減少,絨氈層提前解體,導致營養物質不能運輸到小孢子中去,小孢子不能繼續發育而最終敗育。
[1] 耿三省,王志源,蔣健箴.辣椒雄性不育系小孢子發生的細胞學觀察[J].園藝學報,1994,21(2):165-169.
[2] 譚婷婷,劉立鋒,陳偉.利用分子標記鑒定蘿卜細胞質雄性不育種質的研究[J].山東農業科學,2011(8):8-13.
[3] 馮騁宇.大白菜核基因雄性不育系定向轉育研究[D]. 2006,沈陽:沈陽農業大學.
[4] 張書芳,宋兆華,趙雪云.大白菜細胞核基因互作雄性不育系選育及應用模式[J].園藝學報,1990,17(2):117-125.
[5] VanDerMeerQP,高書國.大白菜的染色體單基因顯性雄性不育性[J].中國蔬菜,1991(5):54.
[6] 魏毓棠,馮輝.大白菜雄性不育遺傳規律的研究[J].沈陽農業大學學報,1992,23(3):260-266.
[7] 馮輝.大白菜核基因雄性不育性的研究[D].沈陽:沈陽農業大學,1996.
[8] 余鳳群,傅廷棟.甘藍型油菜幾個雄性不育系花藥發育的細胞形態學研究[J].武漢植物學研究,1990,8(3):209-216.
(責任編輯:張瑞麟)
S634.1
B
0528-9017(2014)02-0197-03
文獻著錄格式:高宏秀,張瑩.大白菜核不育雄蕊敗育超微結構觀察[J].浙江農業科學,2014(2):197-199,201.
2013-11-08
高宏秀(1980-),江蘇泗陽人,講師,從事生物技術教學與研究工作。E-mail:575963188@qq.com。
張 瑩。
