Kir2.1/Kir2.3 嵌合體的構建與表達
2013-12-07 05:37:54趙志英朱宏謙張國紅
中國藥理學通報 2013年5期
趙志英,朱宏謙,張 璇,劉 麗,張國紅
(1.河北醫科大學藥理學教研室,教育部血管與神經生物學重點實驗室,河北省新藥藥理毒理研究重點實驗室,河北石家莊 050017;2.河北省公安廳國內安全保衛總隊,河北石家莊 050051)
內向整流鉀通道 (inwardly rectifying potassium channel,Kir)的特征是鉀離子容易由細胞外向細胞內流動,表現出強烈的電壓依賴性內向整流特性,這種通道的整流作用有利于維持細胞的靜息電位并參與復極化過程[1]。Kir通道至少由7個家族組成,它們在各種組織中廣泛分布,Kir2.0家族是它的重要一員,主要存在于心臟、泌尿系統、神經系統、骨骼肌等許多組織。Kir2.1和Kir2.3是Kir2.0家族中的重要成員,在心臟,Kir2.1和Kir2.3是心臟內向整流鉀電流的重要組成部分,對維持心肌細胞動作電位起重要作用;在腎臟,Kir2.1和Kir2.3是腎皮質集合管內向整流鉀電流的重要組成部分,對維持腎臟的泌鉀、濾過重吸收等生理功能起重要作用。Kir2.1鉀通道是目前Kir2.0家族中發現的唯一一個與離子通道病有關的通道。編碼Kir2.1鉀通道的基因突變,將會引起安德森綜合征(Andersen’s syndrome,AS),典型的臨床表現為周期性麻痹、室性心動過速性心律失常和明顯的結構異常[2]。研究Kir2.1和Kir2.3的調控機制對研究整個內向整流鉀通道家族的調控機制有重要意義[3]。在研究過程中人們發現Kir2.3可以被細胞內或細胞外的許多信號分子如Mg2+、多胺、H+、蛋白激酶C(PKC)、磷脂酰肌醇4,5二磷酸(PIP2)、花生四烯酸、植物雌激素、ATP等所調控[4-6],而這些調控因素都對 Kir2.1沒有作用或作用很弱,因此我們設想構建不同的Kir2.1和Kir2.3通道嵌合體,為進一步研究Kir2.3的調控機制和通道特異性阻斷劑提供實驗基礎。
1 材料與方法
1.1 主要試劑與儀器 Kir2.3,Kir2.1的cDNA克隆在質粒pGEMHE中。pGEMHE質粒DNA由美國紐約大學西奈山醫學院生理學系Diomedes Logothetis教授饋贈。Pyrobest DNA polymerase、EcoRI、BamHI等限制性核酸內切酶購自日本TaKaRa公司,瓊脂糖凝膠回收試劑盒購自美國Promega公司。基因擴增儀購自德國Biometra公司,Nanoliter Injector RNA注射儀購自美國WPI公司,Geneclamp 500B放大器購自美國Axon公司。
1.2 方法
1.2.1 嵌合體構建 以 N1P3C3為例說明 Kir2.1/Kir2.3嵌合體構建過程。將Kir2.3通道的氨基末端替換為Kir2.1通道的氨基末端,根據GenBank發表的Kir2.1和Kir2.3通道的序列設計出擴增Kir2.1通道氨基末端的引物和擴增Kir2.3通道孔區、跨膜區、羧基末端的引物。Kir2.1-N上游引物包含一個 BamHI酶切位點為:5'-CGGGGATCCATGGGCAGTGTGAGAACC-3';下游引物為:5'-GAGACAAGGAAGGCCGCGGAGAAGATAACCAGCATCCACC-3';以 Kir2.1-pGEMHE質粒DNA為模板,PCR擴增條件為94℃預變性5 min;94℃ 1 min,58℃ 1 min,72℃ 1 min,25 個循環;72℃延伸10 min。Kir2.3-PC上游引物為:5'-GTGGATGCTGGTTATCTTC TCCGCGGCCTTCCTTGTCTCC-3';下游引物包含一個 EcoRI酶切位點為:5'-GATGAATTCGATCCGGGGACCTAGAGAGCCAC-3';以Kir2.3-pGEMHE質粒DNA為模板,PCR擴增條件為94℃預變性5 min;94℃ 1 min,60℃ 1 min,72℃ 1 min,25個循環;72℃延伸10 min。PCR產物用1%瓊脂糖凝膠電泳進行分析,切膠純化,并作為模板進行重疊延伸PCR,擴增條件為94℃預變性5 min;94℃ 1 min,55℃ 1 min,72℃ 1 min,8 個循環;72℃延伸 10 min。再加入 Kir2.1-N上游引物和Kir2.3-PC下游引物進行擴增,擴增條件為94℃預變性 5 min;94℃ 1 min,58℃ 1 min,72℃ 1 min,25 個循環;72℃延伸10 min。PCR產物純化后與pGEMHE質粒DNA進行 EcoRI和 BamHI雙酶切,酶切后進行純化,在T4DNA連接酶作用下進行連接,連接產物轉化TOP10感受態細胞,挑取陽性克隆,擴菌增菌,抽提質粒進行酶切鑒定,送基因測序公司進行測序分析,挑選沒有突變的克隆,命名為N1P3C3。采用同樣的方法構建其它的嵌合體質粒。
1.2.2 嵌合體通道表達 嵌合體通道質粒DNA用限制性核酸內切酶NheI將其線性化,采用RibomaxTMLarge Scale RNA Production Systems-SP6 and T7 Kit試劑盒體外轉錄得到cRNA。分離爪蟾卵母細胞,將cRNA注入爪蟾卵母細胞,在18℃培養1-2天后,用雙電極電壓鉗記錄電流是否表達。灌流液 ND96(mmol·L-1):NaCl 96,KCl 1,MgCl21,CaCl21.8,HEPES 5,用 NaOH 調 pH 為7.4;ND96K(mmol·L-1):KCl 96,NaCl 1,MgCl21,CaCl21.8,HEPES 5,用 KOH 調 pH為7.4。
1.2.3 記錄電流 室溫下(22℃ ~24℃)記錄電流,放大器Mode設為SETUP狀態。按下按鈕Zero V1和Zero V2使兩電極電位顯示為零,用R1和R2測量電極電阻。刺入電極,將放大器Mode調為Voltage clamp狀態,記錄通道電流。信號的采集和分析軟件為pClamp 9.0。實驗所用protocol為:先將膜電位鉗制在0mV,然后從-90 mV逐漸增加(ramp)到+90 mV。
1.2.4 數據處理 采用 Clampfit 9.0(美國Axon公司)和OriginPro 7.0(美國Origin Lab公司),Adobe Illustrator 10等軟件進行圖像處理及數據分析。
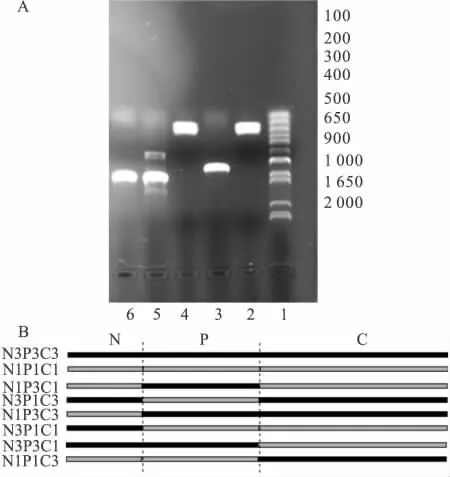
Fig 1 Construction of Kir2.1/Kir2.3 chimeras by over-lap extension PCR
2 結果
2.1 嵌合體構建 由于Kir2.1和Kir2.3通道都屬于同一個Kir2.0亞家族,他們的氨基酸序列具有58%的同源性,因此構建Kir2.1和Kir2.3通道的嵌合體產生功能性通道的可能性很大。根據已知的Kir通道結構,他們以一個較短的氨基末端(N)在細胞內起始,經過兩次跨膜折疊(M1,M2)后,以一個較長的羧基末端(C)在細胞內終止[7]。構建嵌合體通道時首先考慮將通道的氨基和羧基末端進行互換。通道命名原則如下:數字1和3分別代表Kir2.1和Kir2.3通道,字母N代表氨基末端,字母C代表羧基末端,字母P代表兩個跨膜區和孔區。我們首先構建一個嵌合體通道N1P3C3,即通道的氨基末端來源于Kir2.1通道,通道的兩個跨膜區和孔區、羧基末端來源于Kir2.3通道。以Kir2.1質粒DNA為模板,擴增出Kir2.1通道氨基末端,以Kir2.3質粒DNA為模板,擴增出Kir2.3通道跨膜區和孔區、羧基末端,經重疊延伸PCR,擴增產物為N1P3C3,產物長度為1 450 bp,經過膠回收,雙酶切純化后與pGEMHE載體DNA連接,得到N1P3C3-pGEMHE,經測序驗證,挑選沒有堿基突變的克隆。至此,嵌合體通道N1P3C3構建成功(Fig 1 A)。以類似的方法分別將通道的氨基末端、羧基末端、跨膜區和孔區進行互換,成功構建 N3P1C1,N3P3C1,N1P1C3,NIP3C1,N3P1C3 等幾個嵌合體通道。N1P3C3和N3P1C1是將通道的氨基末端進行了互換,N1P3C3是指通道的氨基末端來源于Kir2.1通道,通道的兩個跨膜區和孔區、羧基末端來源于Kir2.3通道;N3P1C1是指通道的氨基末端來源于Kir2.3通道,通道的兩個跨膜區和孔區、羧基末端來源于 Kir2.1通道。N3P3C1和N1P1C3是將通道的羧基末端進行了互換。NIP3C1和N3P1C3是將通道的氨基末端和羧基末端同時進行了互換(Fig 1 B)。
2.2 電壓鉗記錄嵌合體電流 將嵌合體通道N1P3C3表達于卵母細胞,雙電極電壓鉗記錄電流,觀察電流特征。灌流液為ND96K(在此溶液所記錄到的內向電流最大),鉗制電壓為由-90 mV漸變成+90 mV,根據電流隨電壓的變化可得到N1P3C3通道的電壓電流關系曲線,橫坐標上方的電流為外向電流,橫坐標下方的電流為內向電流(Fig 2)。由Fig 2可以看出N1P3C3通道的內向電流明顯大于外向電流,且內向電流與電壓有明顯的線性關系,而外向電流則沒有這種線性關系,反轉電位在ND96K灌流情況下為0 mV左右。Ba2+是鉀通道的阻斷劑,用含 0.3 mmol·L-1BaCl2的ND96K灌流N1P3C3通道。為了更清楚地觀察電流,我們選取觀察高鉀情況下電流幅度較大的-80 mV時的電流,+80 mV的電流為對照,結果顯示Ba2+能夠有效抑制N1P3C3的電流(Fig 3)。這些結果表明N1P3C3的電流特征沒有改變,符合內向整流鉀通道的電流特征。將其它嵌合體通道N3P1C1,N3P3C1,N1P1C3,NIP3C1,N3P1C3 同樣表達于非洲爪蟾卵母細胞,記錄電流,得到了相同的結果,嵌合體通道的電流特性均未改變(結果未顯示)。
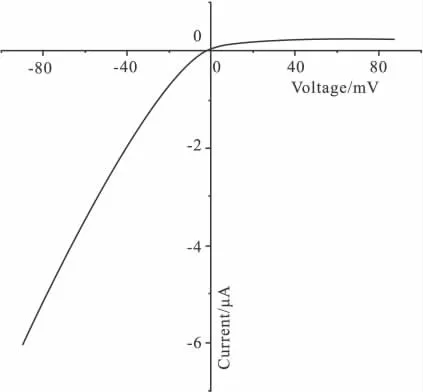
Fig 2 Current-voltage relationship of Kir2.3 currents in high-K+solution(ND96K)
3 討論
內向整流鉀通道在各種組織廣泛分布,其電流特征為鉀離子流入細胞內遠比流出細胞外容易。Kir通道是由4個同源或異源亞基組成的四聚體,每個亞基都是由氨基末端、孔區和跨膜區、羧基末端組成,結構相似,因此構建Kir2.1和Kir2.3的嵌合體通道可行性很高,產生功能性通道的幾率很大[7]。
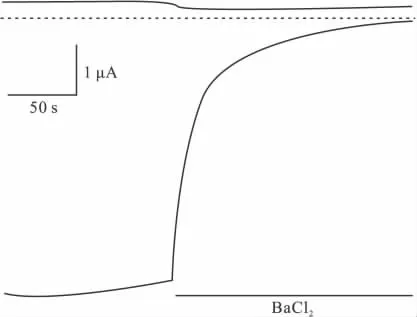
Fig 3 Whole-cell Kir2.3 currents recorded by two electrodes voltage clamp(TEVC)
Kir2.1和Kir2.3是Kir2.0家族重要成員,在神經系統、心血管系統等許多組織都有分布,起著重要的生理作用,許多調控因子對他們都有調節作用。但近年來的研究發現調控因子對Kir2.1和Kir2.3調控作用大相徑庭。PIP2是細胞信號傳導的重要分子[8],PIP2水解使Kir2.3通道電流明顯下降,對Kir2.1通道電流影響不大,但PIP2抑制Kir2.3通道電流的具體機制尚不明確;共同表達Kir2.3通道蛋白與Gβγ亞單位于非洲爪蟾卵母細胞,可以發現Gβγ亞單位能夠完全抑制Kir2.3通道電流,而對Kir2.1通道電流卻沒有作用[9];氫離子在Kir2.3的門控機制中起重要作用,高CO2和低pH值抑制Kir2.3通道電流,對Kir2.1沒有作用;在非洲爪蟾卵母細胞,PKC的激活劑佛波醇酯(PMA)明顯抑制Kir2.3通道電流,對Kir2.1沒有作用[4];花生四烯酸是一種多不飽和脂肪酸,能夠明顯且可逆地增大在中國倉鼠卵巢上皮細胞表達的Kir2.3通道電流,對Kir2.1沒有作用[6];如此可見,許多調控因子對Kir2.3通道電流都有作用,對Kir2.1通道電流沒有作用或作用較弱。Kir2.1和Kir2.3通道同源性很高,而我們的實驗證實,將通道的氨基末端、羧基末端、孔區和跨膜區進行互換,構建的嵌合體通道都產生了功能,電流特性沒有發生改變,這為以后進一步研究調控因子對Kir2.1和Kir2.3通道的作用,確定通道與調控因子的作用位點打下了基礎。先將調控因子作用于嵌合體通道,分析通道的氨基、羧基、孔區和跨膜區哪些片段與調控因子對通道的調節作用有關,在此基礎上分析這些片段上哪些位點的氨基酸是調控因子作用的關鍵位點。
我們對于Kir2.0通道功能的了解主要通過研究人類疾病引起的通道變化、基因敲除小鼠等。由于一直沒有通道特異性阻斷劑,不同亞型Kir2.0通道的研究進展受到了很大限制。已有報道編碼Kir2.1鉀通道的基因突變引起安德森綜合征。但是Kir2.1鉀通道是目前Kir2.0亞家族中發現的唯一一個與離子通道病有關的通道,目前還沒有關于編碼Kir2.3鉀通道的基因突變引起臨床疾病的報道。因此通道特異性阻斷劑的研發必將會成為Kir2.0家族通道生理功能研究、藥物特性研究的里程碑事件。然而,到目前為止,Kir2.0通道的阻斷劑僅限于鋇離子、銫離子,許多非特異性藥物比如:植物雌激素、抗組胺藥、卡維地洛、抗瘧藥甲氟喹、鹽酸米帕林[5,10-13]等都可以明顯抑制 Kir2.3 通道電流,對Kir2.1沒有作用或作用較弱。因此我們可以通過構建Kir2.1和Kir2.3通道的嵌合體,在上述藥物對Kir2.3通道電流特異性抑制作用的基礎上,進一步研究藥物對Kir2.3通道的作用機制,找到它們的作用位點,進而為下一步找到通道的特異性阻斷劑,研究通道的生理病理意義打下基礎。
一些內向整流鉀通道功能改變可能引起某些非遺傳性病理生理變化比如房顫[14,15]。由于Kir2.1和Kir2.3的廣泛分布及其在神經細胞和心肌細胞等的電生理活動中起重要作用,進一步明確Kir2.1和Kir2.3的功能,了解與通道偶聯的受體、第二信使及其他信號分子對Kir2.1和Kir2.3的調控,研究 Kir2.1和 Kir2.3通道的特異性阻斷劑十分重要[15]。而本實驗成功構建的Kir2.1和Kir2.3通道的嵌合體為此打下了堅實的基礎。
[1]Guo D,Ramu Y,Klem A M,Lu Z.Mechanism of Rectification in inward rectifier K+Channels[J].J General Physiol,2003,121(4):261-775.
[2]何海燕,尹永強,李宏杰,等.牛磺酸鎂對缺氧/復氧致大鼠心肌細胞內向整流鉀通道異常的影響[J].中國藥理學通報,2012,28(12):1751-6.
[2]He H Y,Yin Y Q,Li H J,et al.Effects of taurine magnesium coordination compound on abnormal inward rectifier potassium channel current induced by hypoxia reoxygenation in cardiomyocytes of rats[J].Chin Pharmacol Bull,2012,28(12):1751-6.
[3]Kulzer M,Seyler C,Welke F,et al.Inhibition of cardiac Kir2.1-2.3 channels by beta3 adrenoreceptor antagonist SR 59230A[J].Biochem Biophys Res Commun,2012,424(2):315-20.
[4]杜肖娜,何宏濤,王 川,張海林.PKC對兩種表達系統中內向整流性鉀通道調節的不同及機制研究[J].中國藥理學通報,2008,24(12):1615-9.
[4]Du X N,He H T,Wang C,Zhang H L.Study of the mechanism of different regulation of Kir current in two expressions systems by PKC[J].Chin Pharmacol Bull,2008,24(12):1615- 9.
[5]Zhao Z Y,Liu B Y,Zhang G H,et al.Molecular basis for genistein-induced inhibition of Kir2.3 currents[J].Pflugers Arch-Eur J Physiol,2008,456(2):413-23.
[6]Wang C,Mirshahi U L,Liu B Y,et al.Arachdonic acid activates Kir2.3 channels by enhancing channel-PIP2 interactions[J].Mol Pharmacol,2008,73(4):1185-94.
[7]Hibino H,Inanobe A,Furutani K,et al.Inwardly rectifying potassium channels:their structure,function,and physiological roles[J].Physiol Rev,2010,90(1):291-366.
[8]陳興娟,張熙東,張 璇,等.蛋白激酶C和磷脂酰肌醇4,5二磷酸之間相互調節作用的研究進展[J].中國藥理學通報,2012,28(11):1497-9.
[8]Chen X J,Zhang X D,Zhang X,et al.Progress in the research of mutual regulation between PKC and PI(4,5)P2[J].Chin Pharmacol Bull,2012,28(11):1497-9.
[9]Cohen N A,Sha Q,Makhina E N,Inhibition of an inward rectifier potassium channel(Kir2.3)by G-protein betagamma subunits[J].J Biol Chem,1996,271(50):32301-5.
[10]Liu B Y,Jia Z F,Geng X,et al.Selective inhibition of Kir currents by antihistamines[J].Eur J Pharmacol,2007,558(1- 3):21-6.
[11]Ferrer T,Ponce-Balbuena D,Lopez-Izquierdo A,et al.Carvedilol inhibits Kir2.3 channels by interference with PIP2-channel interaction[J].Eur J Pharmacol,2011,668(1-2):72-7.
[12]Lopez-Izquierdo A,Ponce-Balbuena D,Moreno-Galindo E G,et al.The antimalarial drug mefloquine inhibits cardiac inward rectifier K+channels:evidence for interference in PIP2-channel interaction[J].J Cardiovasc Pharmacol,2011,57(4):407-15.
[13]López-Izquierdo A,Aréchiga-Figueroa I A,Moreno-Galindo E G.Mechanisms for Kir channel inhibition by quinacrine:acute pore block of Kir2.x channels and interference in PIP2 interaction with Kir2.x and Kir6.2 channels[J].Pflugers Arch,2011,462(4):505-17.
[14]Ehrlich J R.Inward rectifier potassium currents as a target for atrial fibrillation therapy[J].Cardio-vasc Pharmaco,2008,52(2):129-35.
[15]De Boer T P,Houtman M J,Compier M,et al.The mammalian K(IR)2.x inward rectifier ion channel family:expression pattern and pathophysiology[J].Acta Physiol(Oxf),2010,199(3):243-56.