雙孢蘑菇基質降解能力退化差異表達基因的克隆
2013-11-28 01:39:10陳美元
食藥用菌 2013年1期
關鍵詞:差異
陳美元
(福建省農業科學院食用菌研究所,福州 350014)
雙孢蘑菇(Agaricus bisporus)雜交菌株As2796是福建省蘑菇菌種研究推廣站(現福建省農科院食用菌研究所)選育成功的優良商業蘑菇菌株,多年來一直占據中國雙孢蘑菇栽培面積的80%以上[1]。在該菌株多年的保藏復壯、生產供應的過程中偶然發現了 2株退化突變體 2796-3和2796-5。它們在PDA培養基上生長基本正常,但在糞草培養基、棉籽殼培養基和麥粒培養基上均無法正常生長,在基質特別是多糖類基質的降解能力上發生了嚴重的變異和退化現象。這樣的變異菌種如果流入市場,在 PDA斜面上不斷擴繁成一級母種,而在生產季節卻不能及時制作成二級種和三級種,將造成供種單位和制種戶極大的經濟和信譽損失,并將直接損害廣大菇農的利益。經研究,我們排除了病毒等微生物感染的因素,證明了該退化性狀是基因變異所引起的,并且可以穩定遺傳。
由于 As2796菌株的重要性以及它所具有的雙孢蘑菇商品化雜交菌株的代表性,除了在保藏和供種等方面應加強檢測外,對已發現的該退化現象進行相關基因表達差異及分子機制研究具有重要的理論與實際意義。在前期研究中,我們對雙孢蘑菇 As2796正常菌株及其兩個退化突變體進行了mRNA差異顯示(DDRT-PCR)分析[2],發現有 9個片段存在明顯的上調或下調差異表達,序列分析發現9個差異片段代表了5個基因的差異,它們在雙孢蘑菇基因組數據庫中的代碼分別為 191327、199816、135047、193267、193513。在本研究中,我們對三個菌株中的這5個基因及其上游部分調控序列進行克隆與測序比較,以期獲得這些基因與上游序列在三個菌株中的差異,并進一步探討其可能的分子機制。
1 材料與方法
1.1 試驗材料
(1)菌株。雙孢蘑菇雜交菌株As2796及其退化突變體菌株2796-3、2796-5由福建省農科院食用菌研究所種質資源與遺傳育種研究室保藏并提供。大腸桿菌(Escherichia coli) DH5α購自上海生工生物工程服務有限公司。
(2)試劑。Wizard SV Gel and PCR Clean-up System (DNA 膠純化試劑盒)、pGEM-T Easy Vector System (PCR產物克隆試劑盒)及 SV Minipreps DNA Purification System(質粒提取試劑盒)購自Promega公司,PCR擴增試劑盒、DNA Markers及其它試劑均購自上海生工生物工程服務有限公司。
(3)引物。①差異基因編碼序列擴增引物:根據DDRT-PCR分析中獲得的5個差異基因[2]在雙孢蘑菇轉錄組數據庫中的編碼序列(CDS)設計正反向PCR引物,用于擴增cDNA中的編碼序列,也可擴增DNA中含內含子的編碼序列,引物名稱中的數字與 DDRT-PCR中的差異片段編號相同,且對應基因編號 191327、199816、135047、193267、193513。引物序列見表 1。②差異基因上游調控序列擴增引物:根據DDRT-PCR分析中獲得的5個差異基因[2]在雙孢蘑菇基因組數據庫中的上游序列設計正反向PCR引物,用于擴增DNA中差異基因上游約1 200 bp的調控序列,引物名稱中的數字與DDRT-PCR中的差異片段編號相同,且對應基因編號191327、199816、135047、193267、193513。引物序列見表2。
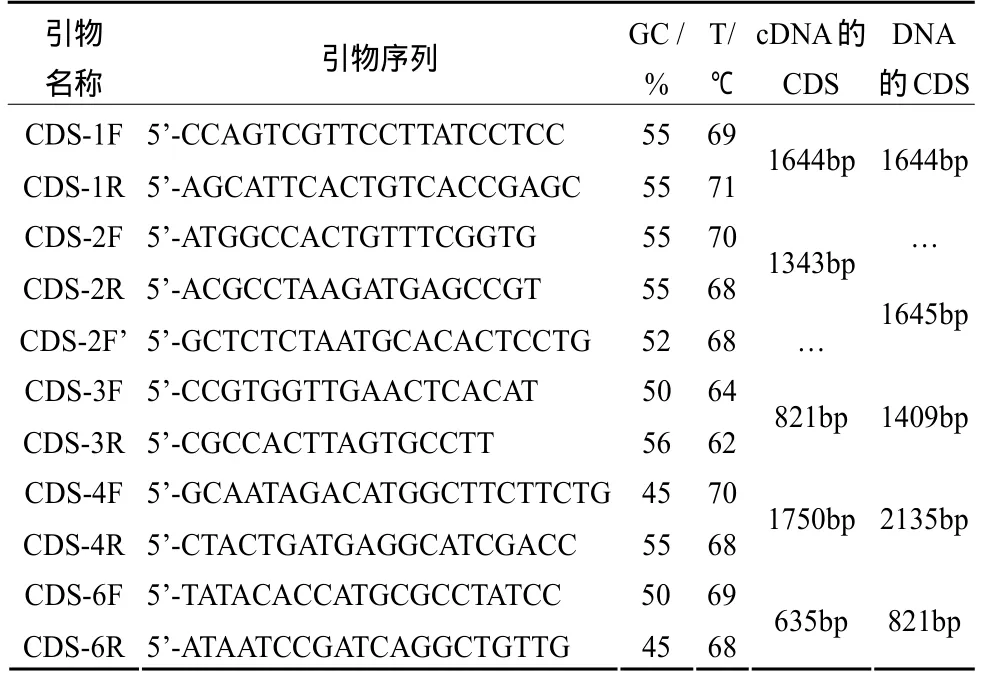
表1 差異基因編碼序列擴增引物
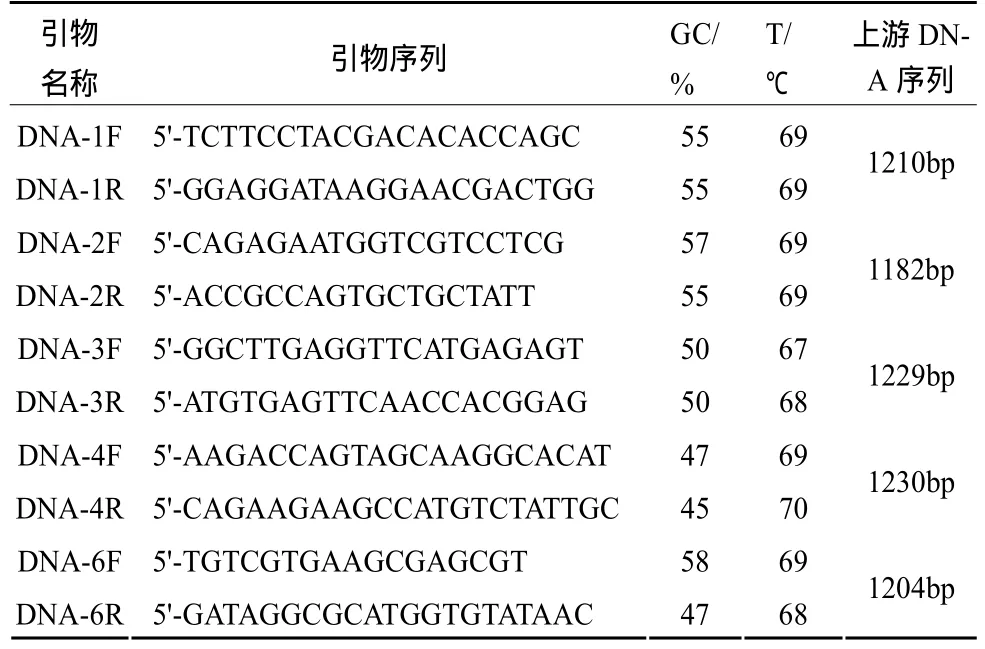
表2 差異基因上游序列擴增引物
1.2 試驗方法
(1)菌絲的培養。菌種接入常規PDA斜面培養基,24 ℃培養三周。
(2)總DNA的提取。按美國Sylvan公司提供的方法并加以改進[3]。
(3)目的 DNA片段的擴增。按文獻[4]的方法稍加改動。總體積20 μL反應體系中含:10×buffer 2 μL;dNTPs (總 2.5 mmol/L)1.5 μL;模板 DNA(15~20 ng/μL)2 μL;正反向特異引物(約 10 μmol/L)各 1 μL;Pfu(0.5U/μL)2 μL;純水10.5 μL。混勻后在臺式離心機上稍加離心,放入PCR儀進行下列循環:94 ℃ 2 min;94 ℃ 1 min,58 ℃ 1 min,72 ℃ 2 min,35 Cycles;72 ℃10 min;4 ℃保持。擴增產物進行1%瓊脂糖凝膠電泳,GeneFinder熒光染料染色拍照。
(4)目的DNA片段的純化、克隆與鑒定。按照 Promega公司 Wizard SV Gel and PCR Clean-up System、pGEM-T Easy Vector System和SV Minipreps DNA Purification System試劑盒說明書進行。
(5)克隆子測序與分析。克隆子質粒DNA送上海生工生物工程技術服務有限公司進行測序,獲得的片段序列用DNAMAN軟件進行序列拼接與序列比對。
2 結果與分析
2.1 正常與退化菌株差異基因及其上游序列的擴增與克隆
以雙孢蘑菇As2796及其2個退化菌株的總DNA為模板,用設計合成的10對特異引物對5個差異基因帶內含子的編碼序列及其上游約 1 200 bp調控序列進行擴增,結果均獲得了目標條帶(圖1,圖2)。割取這些條帶并進行純化,然后克隆到pGEM-T Easy載體中。
2.2 正常與退化菌株差異基因及其上游序列的測序及比較
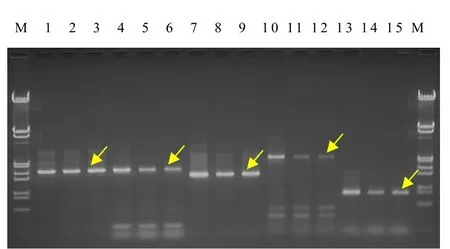
圖1 3個菌株5個差異基因帶內含子CDS的PCR擴增圖
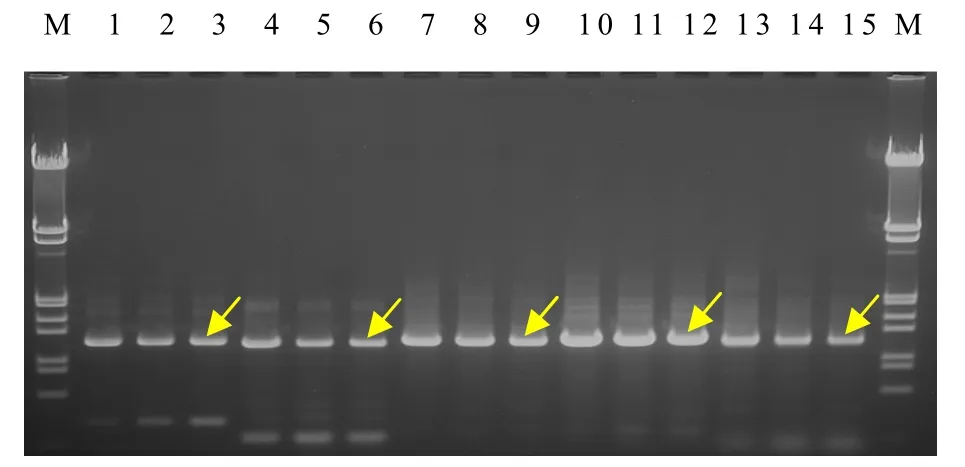
圖2 3個菌株5個差異基因上游序列的PCR擴增圖

圖3 差異基因199816在3個菌株中的序列比較
對克隆子質粒進行測序與序列拼接,比較3個菌株間的序列,結果表明除了差異基因199816在3個菌株中的序列完全一致外(圖3),其余基因與上游序列在三個菌株中均有幾個堿基的差異(圖4)。而這些差異在2個退化菌株中的表現未必一致,重復擴增與測序的結果也顯示這些差異并無明顯的規律性和重復性。因此它們應是PCR擴增及測序過程中產生的誤差,而非實質性差異。

圖4 差異基因135047上游序列在3個菌株中的序列比較
3 討 論
食用菌菌種在保藏、制種及后續的栽培過程中,時有發生長勢變差、出菇延遲、產量下降、品質劣變、抗性減弱等現象,這些現象人們泛稱“退化”。 我國是食用菌生產大國,而食用菌菌種的退化又是一個較為常見的現象,因此國內針對食用菌生產上的退化現象有較多的綜合性報道,內容包括食用菌菌種退化與老化的現象,可能的因素及防止措施。其中提到退化因素可能有遺傳變異、病毒或雜菌污染、無限傳代、營養不良、保藏不當、人工選擇偏差等內外因[5~7]。但在食用菌的退化機理研究方面,除了本實驗室對雙孢蘑菇的叢生變異與基質降解能力退化現象有關于基因變異的初步研究和公開報道[2,8,9],以及Magae等[10]在金針菇和Qiu等[11]在平菇中發現dsRNA病毒感染可導致性狀退化外,未見其他食用菌退化的相關基因與分子機理研究的文獻報道。
我們的研究首次發現一些在雙孢蘑菇基質降解能力退化菌株中差異表達的基因,但這些基因及其上游序列在三個菌株中未能發現可重復的實質性差異。結合DDRT-PCR結果我們判斷正常與退化菌株的這些基因在轉錄上存在量的差異。但結構基因本身及其上游調控序列并未發生實際的變異,退化更可能是影響胞內信號傳導或基因轉錄過程的調控蛋白基因變異引起的。這為進一步深入研究雙孢蘑菇分解基質能力退化的分子機理奠定了基礎。
[1]王澤生,廖劍華,陳美元.我國雙孢蘑菇育種研究與產業發展[J].食用菌學報,2010,(增刊):19-20.
[2]陳美元,王澤生,廖劍華,等.雙孢蘑菇基質降解能力退化的 DDRT-PCR分析[J].菌物學報,2010,29(5):707-712.
[3]陳美元,廖劍華,盧政輝,等.雙孢蘑菇栽培菌株遺傳多樣性的DNA指紋分析[J].菌物學報,2007,26(增刊):128-137.
[4]陳美元,廖劍華,王波,等.中國野生蘑菇屬90個菌株遺傳多樣性的DNA指紋分析[J].食用菌學報,2009,16(1):11-16.
[5]丁湖廣.菌種退化與老化原因及防止措施[J].特種經濟動植物,2006,1:39-40.
[6]劉海英,董月香,周廷斌,等.食用菌菌種的退化及復壯[J].食用菌,2003,25(6):16-17.
[7]Fritsche.G..Maintenance Rejuvenation and Improvement of HORST-U1[J].Genetic and Breeding of Agaricus,1991:145-152.
[8]陳美元,王澤生,廖劍華,等.雙孢蘑菇叢生變異的RAPD分析及差異DNA片段的克隆[J].廈門大學學報(自然科學版),2003,42(5):657-660.
[9]陳美元,王澤生,廖劍華,等.雙孢蘑菇基質降解能力退化的差異蛋白質組學分析[J].菌物學報,2011,30(3):508-513.
[10]Magae Y,Hayashi N.Double-stranded RNA and virus-like particles in the edible basidiomycete Flammulina velutipes (Enokitake) [J].FEMS Microbiol Lett.,1999, 180(2):331-5.
[11]Qiu L, Li Y, Liu Y, et al.. Particle and naked RNA mycoviruses in industrially cultivated mushroom Pleurotus ostreatus in China[J]. Fungal Bio., 2010,114(5-6): 507-13.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
