傳統海產調味品中微生物及其發酵作用研究進展*
2013-11-21 10:02:02李瑩白鳳翎勵建榮
食品與發酵工業 2013年10期
關鍵詞:研究
李瑩,白鳳翎,勵建榮
(渤海大學化學化工與食品安全學院,遼寧省食品安全重點實驗室,“食品貯藏加工及質量安全控制工程技術研究中心”遼寧省高校重大科技平臺,遼寧錦州,121013)
傳統發酵海產調味品通常以新鮮的魚或蝦為原料,添加高濃度的鹽(一般為20% ~30%),經過1~2年的長時間發酵,利用原料自身蛋白酶、細菌蛋白酶及各種酶類分解蛋白質、脂類、碳水化合物大分子物質形成氨基酸、脂肪酸及各種風味物質,賦予產品鮮、香、咸等品質特征。此外,產品中富含有易于人體吸收的氨基酸、多肽、維生素以及鈣磷鐵等礦物質,具有提高免疫力,防貧血,降血壓血脂等功效。
世界很多國家生產各種傳統發酵海產調味品,如中國的魚露、蝦油蝦醬,韓國的 Jeotkal、Sikhae,日本的Nakazuke,印度尼西亞的 Bakasang,加納的Momoni,埃及的 Feseekh,伊朗的 Mahyaveh,泰國的 Plaasom以及Kungchao等等。在東南亞國家,傳統發酵海產調味品是當地居民飲食中十分重要的佐餐食品,可作為烹飪的調味劑和菜肴的增香劑,有時甚至被作為下飯的一道小菜[1]。不同國家海產調味品在生產方法、原料配方和消費者的喜好等方面都存在一定的差異[2]。本文從發酵微生物對產品品質作用與影響的角度出發,對各種傳統發酵海產調味品中微生物的構成、微生物在發酵過程中的功能特性的研究進展進行綜述,旨在利用微生物的發酵作用改善產品風味,降低產品鹽度,縮短發酵周期,提高經濟效益。
1 傳統發酵海產調味品中微生物菌相構成
不同發酵海產品在發酵過程中微生物的種類不同,同一產品在不同發酵時期微生物的種類也不同。Sanchez和Klitsaneephaiboon對傳統發酵魚醬油Patis發酵相關的微生物進行研究,結果表明發酵初期的優勢菌是芽孢桿菌屬的短小芽孢桿菌、凝結芽孢桿菌和枯草芽孢桿菌,發酵中期優勢菌為地衣芽孢桿菌、生皺微球菌和表皮葡萄球菌,發酵終期優勢菌為變異微球菌和腐生葡萄球菌[6]。
2 傳統海產調味品中微生物的發酵作用
針對傳統發酵海產調味品中微生物的研究主要集中在特征性嗜鹽菌的篩選和分類鑒定方面,而對嗜鹽菌的發酵作用研究相對偏少,研究主要包括耐鹽蛋白酶的功能、香氣和風味物質形成和生物胺降解等方面。
2.1 微生物源耐鹽蛋白酶
傳統發酵海產調味品的生物降解過程主要是將不溶的蛋白質轉化為可溶的或懸浮的多肽和氨基酸,所依賴的是海產自身的和來自嗜鹽菌的蛋白酶。在發酵中后期,嗜鹽菌的耐鹽蛋白酶的作用更為突出。
現今商用微生物源蛋白酶大多來自嗜溫型微生物,一般在中性至弱堿性pH值范圍、常溫和低離子強度條件下顯示高蛋白酶活性,而在極端條件下活性較差。如地衣芽孢桿菌蛋白酶在pH 4.0時活性較低,隨NaCl濃度的增加活性逐漸降低[7]。相反,極端環境微生物產生的蛋白酶卻有不同的表現,一些古細菌蛋白酶在5.8% ~30%NaCl條件下均具有活性,如亞洲嗜鹽堿桿菌172P1產生的糜蛋白酶在30%NaCl下具有最佳活性[8]。
傳統發酵海產調味品中的極端嗜鹽菌具有形成蛋白酶的能力,如來自泰國魚醬的4株嗜鹽四聯球菌M11、MS33、MRC5-5-2和 MRC10-7-8在高鹽濃度下對魚肉蛋白都具有分解活性,所產生的寡肽和低聚肽是形成產品風味物質的前體[3]。
嗜鹽菌蛋白酶在較低鹽度條件下活性很低,在一定范圍內酶活性隨著鹽濃度的增高逐漸增強。來自泰國鳳尾魚魚醬油的嗜鹽菌菌株Virgibacillus sp.SK33產生耐鹽絲氨酸蛋白酶,其活性在低鹽度條件下較弱,隨鹽濃度的增加活性逐漸增強,在25%NaCl時活性最高。相比而言,這種天然蛋白酶比堿性蛋白酶和復合蛋白酶具有更好的水解活性[9]。
2.2 微生物源性香味和風味物質
微生物發酵作用對傳統發酵海產調味品特征品質形成具有非常重要的呈獻作用,一是依靠酶解產物或產物之間相互作用形成的醛、酮、醇、酯類等香氣成分,二是來自酶解作用形成的氨基酸、寡肽類等風味物質[10]。Jirawat等將 T.halophilus MS33 和 MRC5-5-2接種至25%NaCl的魚肉湯中,檢測出形成了1-丙醇、2-甲基丙醛和苯甲醛等揮發性物質,與自然發酵魚醬中的主要揮發性物質組成基本一致[3]。由此說明傳統發酵海產調味品中的微生物對風味和香氣物質的形成起著十分重要的作用。
2.2.1 香味物質
兩組均行鼻內鏡手術治療。常規局麻,觀察組2例,對照組1例因病變范圍廣泛行低血壓控制性全麻。采用Messerk-linger術式,切除鼻腔內鉤突及息肉,清除不可逆病變組織,開放病變鼻竇;術后采用藻酸鈣敷料填塞止血,48h后取出。
傳統發酵海產調味品有其特有的香味,如氨味、芝士味、肉香味和奶油香味。氨味揮發性物質包括氨、胺和含氮化合物,它們對產品的香味有增強作用。芝士味主要是由酮類物質形成的,而醛類物質呈現肉香味,小分子量的脂肪酸、乙酸和n-丁酸等呈現奶油香味。
大量研究發現,海產調味品中微生物參與香味物質的形成。當腌魚調料含有抗生素或防腐劑時,產品就不具有典型的芳香氣味,同時揮發性脂肪酸(VFA)的含量很低。VFA不是脂肪酸自然氧化的產物,而是來自于嗜鹽菌的次級代謝產物。嗜鹽菌代謝形成的氨基酸、寡肽進一步分解產生的醇類、酮類、醛類以及酸類成分是香味揮發性物質的來源。例如亮氨酸分解形成的代謝產物2-甲基丁醛、3-甲基丁醛和3-甲基丁醇主要是芝士風味及肉香味的揮發性成分[11]。
Cha等從傳統發酵魚醬、蝦醬中檢測出多達155種揮發性物質,其中占絕大多數為醛、酮、醇、酯類、芳烴、含氮和含硫化合物等揮發性物質,一些特征性含氮雜環化合物如吡嗪主要存在蝦醬中[12]。泰國魚醬中的嗜鹽乳酸菌有助于醇類的積累,同時也產生少量的乙酸乙酯和不同含量的丙酮、2-丁酮、2,3-丁二酮和環己酮等揮發性成分,其中,菌株 T.halophilus MS33、MRC5-5-2、MCD10-5-15 有利于 2-甲基丙醛的形成,菌株MRC10-1-3有利于苯丙醛形成[3]。
發酵海產調味品中微生物發酵過程中在形成各種香氣物質的同時,也產生不受人們歡迎的二甲基硫醚、二甲基二硫醚、二甲基三硫醚等臭味物質。一些細菌具有抑制這些物質形成的能力,Fukami等人的研究表明,在魚醬中添加木糖葡萄球菌(Staphylococcus xylosus)后,終產品中的二甲基二硫醚、二甲基三硫醚含量顯著減少[13]。
2.2.2 風味物質
傳統發酵海產調味品最主要的風味是由揮發性化合物產生的咸味,其次是由氨基酸、寡肽和多肽呈現的特殊風味。海產調味品的咸味主要是由氯化鈉和一些其他無機鹽以及谷氨酸單鈉鹽、天門冬氨酸單鈉鹽產生的。例如,蠔油中呈鮮味成分有琥珀酸鈉、谷氨酸鈉等,呈甜味成分包括甘氨酸、丙氨酸、脯氨酸等[14]。Saisithi等發現在泰國腌魚調料Nam-pla中的呈鮮物質主要含有組氨酸、酪氨酸和色氨酸等[15]。泰國魚醬中的T.halophilus發酵魚肉蛋白產生的氨基酸主要是組氨酸,也形成微量的酪氨酸[3]。
不同海產調味品中微生物的發酵作用與風味形成具有密切的聯系,Beddows等對馬來西亞Budu的研究結果也表明微生物對風味物質以及VFA的形成作用很大[16]。
2.3 微生物降解生物胺作用
傳統海產調味品發酵過程會產生生物胺,過量生物胺會使發酵食品產生嚴重的食品安全問題[17-20]。傳統發酵海產調味品中生物胺的主要成分組胺是由多種微生物的組氨酸脫羧酶誘導產生的,魚醬油中含有超過1 000 mg/L 的組胺[21-24]。在發酵過程中,可以利用微生物控制組胺的形成,提高產品安全性[25]。研究表明,一些嗜鹽菌如枝芽孢桿菌屬、極端嗜鹽古細菌等具有降解生物胺形成的作用。一般是通過自身組胺氧化酶氧化降解組胺為氨、醛以及過氧化氫的途徑,減少在食品內的累積[26]。魚醬油中的嗜鹽微生物Brevibacillus sp.SK35可以使組胺降解率達99%[27]。Zaman 等的研究表明,Staphylococcus carnosus FS19和Bacillus amyloliquefaciens FS05分別使魚醬油中組胺含量降低了27.7%和15.4%[28]。Dapkevicius等從魚露中分離的78株奧默柯達酵母菌(Kodamaea ohmeri)中有胺氧化酶活性的有48株,能降解魚醬中50%的組胺的菌有2株[29]。
3 傳統海產調味品中微生物的發酵作用研究狀況
世界范圍內,對傳統發酵海產品中微生物學研究主要包括兩個方面,一是對發酵過程和產品中微生物形態學和分類學研究,二是對微生物和產品品質之間的相互作用進行研究,目前已從最初的嗜鹽菌的分離鑒定及分類學研究逐步向產品的發酵作用與功能方面深入。
表1是近些年傳統海產調味品中微生物的發酵作用及功能研究狀況,從中可以看出微生物的作用和功能主要包括四個方面:一是研究微生物的蛋白酶作用與功能以及對產品品質的影響;二是研究微生物形成產品的風味物質作用和能力;三是研究微生物降解組胺的作用;四是以微生物作為發酵劑提高產品品質和功能方面的研究。
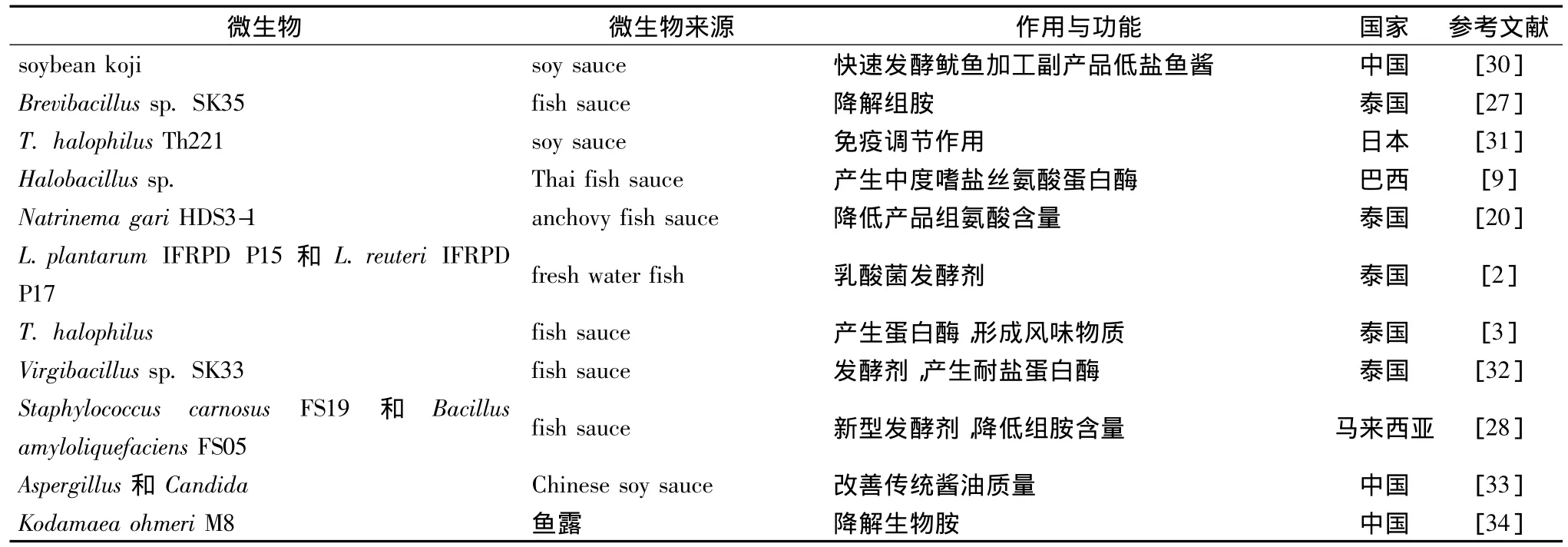
表1 傳統發酵海產調味品中微生物的發酵作用及功能研究Table 1 Fermentation and function of microorganism from traditional fermented seafood products
從研究傳統發酵海產調味品中微生物的區域來看,主要有泰國、馬來西亞、菲律賓、印度尼西亞等東南亞國際以及中國、韓國和日本等東亞國家,也包括加納、埃及和巴西等非亞洲國家。研究比較廣泛和深入的國家首選泰國,泰國海產調味品種類眾多,包括Plaa-som、Kungchao、Nam-pla 和 Pla-ra 等[35],但海產調味品中微生物的種類和發酵作用功能方面差異性很大。例如Halobacillus sp.、T.halophilus和Virgibacillus sp.SK33三種嗜鹽菌均可形成耐鹽蛋白酶,其中T.halophilus主要對魚醬產品的風味形成發揮主導作用,Virgibacillus sp.SK33的主要作用是加速發酵進程,縮短發酵周期。從魚醬油和 anchovy fish sauce產品中分離的菌株 Brevibacillus sp.SK35和Natrinema gari HDS3-1可以有效地降低組氨酸含量,從fresh water fish中分離的L.plantarum IFRPD P15和L.reuteri IFRPD P17也作為發酵劑應用于縮短發酵周期和改善海產調味品質量方面。
馬來西亞對海產調味品的微生物學研究主要是利用Staphylococcus carnosus FS19和Bacillus amyloliquefaciens FS05作為Budu和Bakasang等的發酵劑應用于生產實踐中,降低產品中組胺的含量。在菲律賓魚醬Tinabal和Balbakwa的生產中,微生物發酵作用主要在于形成濃郁的風味物質[36]。
韓國和日本的研究主要是在產品和發酵過程中分離特征性的微生物。例如,從韓國傳統發酵海產品中分離得到的微生物,經鑒定包括從鹽腌制的北美烏魚中分離的Halomonas jeotgali sp.nov.以及腌蝦中分離的 Natronococcus jeotgali sp.nov.[37-38]。在日本soy sauce moromi中分離出具有免疫調節作用的菌株Tetragenococcus halophilus Th221。
在我國,目前應用于發酵海產調味品中的研究主要還處在初級階段,從產品和發酵過程中分離特征性嗜鹽菌。在應用方面,以Kodamaea ohmeri M8菌株在25%NaCl時添加到魚露發酵過程中,使組胺和酪胺的降解率分別達到64.4% 和72.4%[34]。
4 結論與展望
傳統發酵海產調味品是各國人民智慧的結晶,長期以來,由于對發酵過程中微生物作用的認知不足,致使產品的品質和安全性的提升徘徊不前。然而,近些年經過各國微生物工作者的潛心研究,已經獲得各種傳統海產調味品中一定量的微生物學信息,同時也開展了微生物對蛋白質的降解作用、產品風味與香氣物質形成和生物胺降解的作用和功能,進而利用特征性微生物菌株作為發酵劑改善產品品質和提升食品安全性應用研究。如此,對傳統發酵海產調味品微生物學研究進入一個新的時期。然而,人們對傳統發酵海產品極端環境下微生物的菌相構成、生存機制、發酵作用與功能認知才剛剛開始,未知的還很多,有待于科研工作者不懈的努力,對高鹽環境下微生物的分布狀況及與產品之間的相互作用關系進行分析與研究。從機理上探知微生物對產品品質的作用和影響,從應用上獲取特征性微生物應用到發酵過程中,以降低鹽度、增強風味、提高安全性為指標,達到縮短發酵周期,提高產品品質和經濟效益的目的。
[1]Mehdi Z,Hossein N,Mohammad H E,et al.Chemical and microbial properties of mahyaveh,a traditional Iranian fish sauce[J].Food Control,2012,23(2):511-514.
[2]Saithong P,Panthavee W,Boonyaratanakornkit M,et al.Use of a starter culture of lactic acid bacteria in plaa-som,a Thai fermented fish[J].Journal of Bioscience and Bioengineering,2010,110(5):553-557.
[3]Udomsil N,Rodtong S,Tanasupawat S,et al.Proteinaseproducing halophilic lactic acid bacteria isolated from fish sauce fermentation and their ability to produce volatile compounds[J].International Journal of Food Microbiology,2010,141(3):186-194.
[4]Satomi M,Kimura B,Mizoi M,et al.Tetragenococcus muriaticus sp.nov.,a new moderately halophilic lactic acid bacterium isolated from fermented squid liver sauce[J].International Journal of Systematic Bacteriology,1997,47(3):832-836.
[5]Antonio V,Joaquin J N,Aharon O.Biology of Moderately Halophilic Aerobic Bacteria[J].Microbiology and Molecular Biology Reviews,1998,62(2):504-544.
[6]Sanchez P C,Klitsaneephaiboon M.Traditional fish sauce(patis)fermentation in the Philippines[J].Philippine Agriculturist,1983,66(3):69-251.
[7]Gupta A,Roy I,Patel RK,et al.One-step purification and characterization of an alkaline protease from haloalkaliphilic Bacillus sp.[J].Journal of Chromatography A,2005,1075(1-2):103-108.
[8]Kamekura M,Seno Y.A halophilic extracellular protease from a Halophilic archaebacterium strain 172 P1[J].Biochemistry and Cell Biology,1990,68(1):352-359.
[9]Sinsuwan S,Rodtong S,Yongsawatdigul J.A NaCl-stable serine proteinase from Virgibacillus sp.SK33 isolated from Thai fish sauce[J].Food Chemistry,2010,119(2):573-579.
[10]Christensen J E,Dudley E G,Pederson J A,et al.Peptidases and amino acid catabolism in lactic acid bacteria[J].Antonie van Leeuwenhoek,1999,76(1-4):217-246.
[11]Masson F,Hinrichsen L,Talon R,et al.Factors influencing leucine catabolism by a strain of Staphylococcus carnosus[J].International Journal of Food Microbiology,1999,49(3):173-178.
[12]Cha Y J,Cadwallader K R.Volatile components in saltfermented fish and shrimp pastes[J].Journal of Food Science,1995,60(1):19-24.
[13]Fukami K,Funatsu Y,Kawasaki K,et al.Improvement of fish sauce odor by treatment with bacteria isolated from the fish-sauce mush(moromi)made from frigate mackerel[J].Journal of Food Science,2004,69(2):45-49.
[14]綦翠華.值得開發的海產鮮味調味品[J].中國調味品,2006(1):85-90.
[15]Saisithi P,Kasemsarn B O,Liston J,et al.Microbiology and chemistry of fermented fish[J].Journal of Food Science,1966,31(1):105-110.
[16]Beddows C G,Ardeshir A G,Daud W J B.Development and origin of the volatile fatty acids in Budu[J].Journal of Science of Food and Agriculture,1980,31(1):86-92.
[17]Lonvaud-Funel A.Biogenic amines in wines:Role of lactic acid bacteria[J].FEMS Microbiology Letters,2001,199(1):9-13.
[18]Maintz L,Novak N.Histamine and histamine intolerance[J].The American of Clinical Nutrition,2007,85(5):1 185-1 196.
[19]Til H P,Falke H E,Prinsen M K,et al.Acute and subacutetoxicity of tyramine,spermidine,spermine,putrescine and cadaverine in rats[J].Food and Chemical Toxicology,1997,35(3-4):337-348.
[20]Tapingkae W,Tanasupawat S,Parkin K L,et al.Degradation of histamine by extremely halophilic archaea isolated from high salt-fermented fishery products[J].Enzyme and Microbial Technology,2010,46(2):92-99.
[21]Santos M H.Biogenic amines:their importance in foods[J].International Journal of Food Microbiology,1996,29(2):213-231.
[22]Haas H L,Sergeeva O A,Selbach O.Histamine in the nervous system[J].Physiological Reviews,2008,88(3):1183-1241.
[23]Michihata T,Kato D,Yano T,et al.Contents of polyamines in ISHIRU(fish sauce)[J].Journal of the Japanese Society for Food Science and Technology,2006,53(6):337-343.
[24]Kuda T,Miyawaki M.Reduction of histamine in fish sauces by rice bran nuka[J].Food Control,2010,21(10):1322-1326.
[25]Canadian Food Inspection Agency(CFIA).Fish Products Standard and Methods Manual[S].Guide to Food Labelling and Advertising,2009.
[26]Garcia-Ruiz A,González-Rompinelli E M,Bartolomé B,et al.Potential of wine associated lactic acid bacteria to degrade biogenic amines[J].International Journal of Food Microbiology,2011,148(2):115-120.
[27]Sinsuwan S,Montriwong A,Rodtong S,et al.Biogenic amines degradation by moderate halophile,Brevibacillus sp.SK35[J].Journal of Biotechnology,2010,150(Supplement):316.
[28]Zaman M Z,Abu Bakar F,Jinap S,et al.Novel starter cultures to inhibit biogenic amines accumulation during fish sauce fermentation[J].International Journal of Food Microbiology,2011,145(1):84-91.
[29]Dapkevicius M L N,Nout M J,Rombouts F M,et al.Biogenic amine formation and degradation by potential fish silage starter microorganisms[J].International Journal of Food Microbiology,2000,57(1):107-114.
[30]Xu W,Yu G,Xue C,et al.Biochemical changes associated with fast fermentation of squid processing by-products for low salt fish sauce[J].Food Chemistry,2008,107(4):1597-1604.
[31]Masuda S,Yamaguchi H,Kurokawa T,et al.Immunomodulatory effect of halophilic lactic acid bacterium Tetragenococcus halophilus Th221 from soy sauce moromi grown in high-salt medium[J].International Journal of Food Microbiology,2008,121(3):245-252.
[32]Sinsuwan S,Nawong S,Rodtong S,et al.Characterization of Virgibacillus sp.SK33 cell-bound proteinases and its application as a starter culture for fish sauce fermentation[J].Abstracts/Journal of Biotechnology,2008,136(Supplement):S717-S742.
[33]Yan Y,Qian Y,Ji F,et al.Microbial composition during Chinese soy sauce koji-making based on culture dependent and independent methods[J].Food Microbiology,2013,34(1):189-195.
[34]楊利昆,付湘晉,胡葉碧等.魚露中生物胺降解菌的篩選及其特性[J].食品科學,2012,33(11):158-162.
[35]Rhee S J,Lee J E,Lee C H.Importance of lactic acid bacteria in Asian fermented foods[J].Microbial Cell Factories,2011,10(1):S5.
[36]Van Veen A G.In:fish as food(ed.G.Borgstrom).Fermented and dried seafood products in Southeast Asia[M].New York:Academic press,1965:227-250.
[37]Kim M S,Roh S W,Bae J W.Halomonas jeotgali sp.nov.,a new moderate halophilic bacterium isolated from a traditional fermented seafood[J].The Journal of Microbiology,2010,48(3):404-410.
[38]Roh S W,Nam Y D,Chang H W,et al.Natronococcus jeotgali sp.nov.,a halophilic archaeon isolated from shrimp jeotgal,a traditional fermented seafood from Korea[J].International Journal of Systematic and Evolutionary Microbiology,2007,57(9):2 129-2 131.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
