擬南芥prr5突變體對ABA的響應(yīng)*
2013-11-10 11:14:40趙小英劉選明朱詠華
激光生物學(xué)報 2013年6期
關(guān)鍵詞:生長
向 芬,黃 帥,趙小英,劉選明,朱詠華*
(1.湖南大學(xué)生物學(xué)院,植物基因組學(xué)與發(fā)育調(diào)控湖南省重點實驗室,湖南長沙 410082;2.湖南省茶葉研究所,湖南長沙 410125)
植物激素脫落酸 (Abscisic acid,ABA)調(diào)控植物生長和發(fā)育的許多方面,例如:促進種子成熟、休眠,抑制種子胚的萌發(fā)、植株根的伸長和葉片氣孔的關(guān)閉等等[1,2]。此外,脫落酸還是一種重要的抗逆誘導(dǎo)因子,介導(dǎo)植物對逆境如干旱、鹽堿、低溫等生理反應(yīng),因此對脫落酸信號轉(zhuǎn)導(dǎo)進行研究對植物生長的調(diào)控具有重要的意義。近年來,與脫落酸信號轉(zhuǎn)導(dǎo)有關(guān)的中間體雖然陸續(xù)被揭曉,但對脫落酸的信號轉(zhuǎn)導(dǎo)機制了解還較少。
在高等植物中,生物鐘通過振蕩器的作用來調(diào)節(jié)下游功能基因的表達,達到調(diào)控許多錯綜復(fù)雜的新陳代謝和壓力響應(yīng)網(wǎng)絡(luò)的目的[3]。PRRs(pseudoresponse regulators)家族中的 PRR7、PRR9基因是生物鐘振蕩器的核心成員[4,5]。有研究發(fā)現(xiàn),植物信號網(wǎng)絡(luò)包括ABA信號轉(zhuǎn)導(dǎo)途徑受內(nèi)源生物鐘節(jié)律調(diào)節(jié),受ABA信號轉(zhuǎn)導(dǎo)途徑調(diào)控的冷脅迫、氣孔運動同時也受生物鐘的調(diào)控[6]。有研究表明,在野生型植株中ABA含量黎明時較低,在中午12點增加了2倍,但是在prr9/7/5三缺失突變體中ABA含量一直很高,說明PRR9/7/5抑制了生物合成ABA途徑[3]。因此,生物鐘振蕩器的核心成員PRR7、PRR9與同是PRRs家族的PRR5可能為ABA信號轉(zhuǎn)導(dǎo)途徑的相關(guān)基因。
為了探討PRR5基因是否為ABA信號轉(zhuǎn)導(dǎo)途徑的相關(guān)基因。本研究比較分析了外源ABA對prr5突變體種子萌發(fā)、根長生長的影響,以及外源ABA對PRR5基因表達的調(diào)節(jié),并驗證了NaCl對prr5突變體種子萌發(fā)的影響,進一步闡明PRR5基因在擬南芥中的功能。
1 材料與方法
1.1 材料
擬南芥[Arabidopsis thaliana(L.)Heynh.]哥倫比亞生態(tài)型野生型(WT)Col-0為湖南大學(xué)生物學(xué)院植物分子生物學(xué)實驗室保存。Col-0背景的prr5缺失突變體純合子由美國加州大學(xué)洛杉磯分校林辰濤實驗室贈送。擬南芥種子用70%乙醇滅菌30 s,10%次氯酸鈉滅菌10 min,無菌水沖洗4~5次,于黑暗下4℃春化4 d后,再用0.1%的瓊脂懸液懸浮種子后點播在含0.8%瓊脂的MS固體培養(yǎng)基上,置于22℃全日照光照培養(yǎng)箱中培養(yǎng)[7]。
1.2 方法
1.2.1 ABA處理 種子萌發(fā)試驗,把春化后的野生型和 prr5突變體種子點播于不同濃度(0、0.3、0.6、0.9、1.2μmol·L-1)ABA、不同濃度(0、50、100、150 mmol·L-1)NaCl的MS固體培養(yǎng)基上,連續(xù)7 d觀察統(tǒng)計各種子的萌發(fā)及萌發(fā)后生長情況[8,9]。根長試驗,把在MS固體培養(yǎng)基上生長6 d的幼苗轉(zhuǎn)移到含不同 ABA 濃度(0、0.1、1、10、50μmol·L-1)的MS固體培養(yǎng)基上生長6 d后再測量根長。數(shù)據(jù)作圖采用Microsoft Excel 2003,數(shù)據(jù)分析采用 DPS v7.55系統(tǒng)進行統(tǒng)計分析。表達分析處理試驗,取生長7 d的擬南芥Col-0幼苗,根部分別置于含不同濃度(0、1、10、50、100μmol·L-1)ABA 的 MS 液體培養(yǎng)基中處理4 h,上述處理后的幼苗以液氮速凍,保存于-80℃冰箱中用于目標(biāo)基因的表達分析[8]。
1.2.2 RNA 提取 按Easy Way RNA Mini Kit試劑盒(安比奧生物技術(shù)有限公司)說明書提取不同濃度ABA處理過的Col-0幼苗總RNA,按照Invitrogen M-MLV Reverse Transcriptase Instruction說明書進行逆轉(zhuǎn)錄,合成 cDNA[10]。該合成鏈作為 RT-PCR的模板,用于檢測PRR5基因的表達水平。引物序列見表1。
1.2.3 實時熒光定量 PCR(real time quantitative PCR)反應(yīng)體系 RT-PCR以上述合成的cDNA為模板,反應(yīng)體系為 20μL,包括雙蒸水 10.8μL,10 × 緩沖 液 2μL,10 mmol·L-1dNTPs 0.4μL,MgCl21.6μL,引物各 0.1μL,SYBR Green I 0.5μL,ROX 0.1μL,2.5 U·μL-1Jump Start Taq 聚合酶(Sigma)0.4μL,cDNA 模板 4μL。其中,ROX 為被動參考染料。反應(yīng)程序為95℃ 10 min,95℃ 30 s,58℃ 30 s,72℃ 30 s,40個循環(huán)。試驗數(shù)據(jù)采用 MxPro軟件(Stratagene)進行分析[11]。ACT2的表達水平被用作均一化的內(nèi)參,未經(jīng)ABA處理過的樣本作為參比樣本,檢測樣品均為3次重復(fù)。
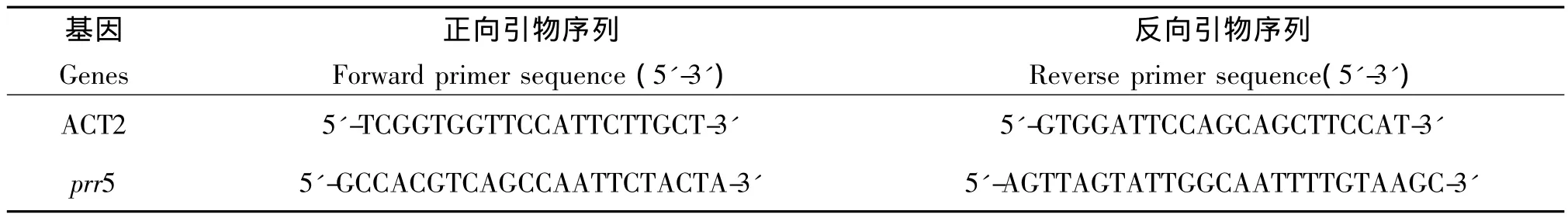
表1 實時定量PCR引物序列Tab.1 Primer sequences used in real time quantitative PCR analysis
2 結(jié)果和分析
2.1 ABA處理對prr5突變體種子萌發(fā)的影響
ABA抑制種子的萌發(fā),為了初步確定prr5突變體是否與ABA信號調(diào)控關(guān)聯(lián),將野生型與prr5突變體播種在含不同濃度(0、0.3、0.6、0.9、1.2μmol·L-1)ABA的MS培養(yǎng)基中,培養(yǎng)7 d后對種子萌發(fā)率及子葉變綠率進行統(tǒng)計分析,圖1顯示,經(jīng)不同濃度ABA處理7 d后,prr5種子萌發(fā)率較野生型高,除0.3μmol·L-1濃度呈顯著差異,其他濃度都呈極顯著差異,即表示prr5對ABA弱敏感。prr5突變體子葉變綠率在 ABA濃度為0.3μmol·L-1以下比野生型高。同時,將prr5突變體種子點播于含不同濃度(0.3、0.9μmol·L-1)ABA 的 MS 固體培養(yǎng)基上,連續(xù)5 d觀察統(tǒng)計種子的萌發(fā)情況。從圖2可以看出,prr5 種子的萌發(fā)率在 ABA 濃度為0.3、0.9μmol·L-1時一直較野生型高,其中,在 ABA濃度為0.3μmol·L-1的MS培養(yǎng)基上突變體萌發(fā)率呈顯著或極顯著增加,在 ABA 濃度為 0.9μmol·L-1上第 2、3 d 其萌發(fā)率都存在極顯著差異。
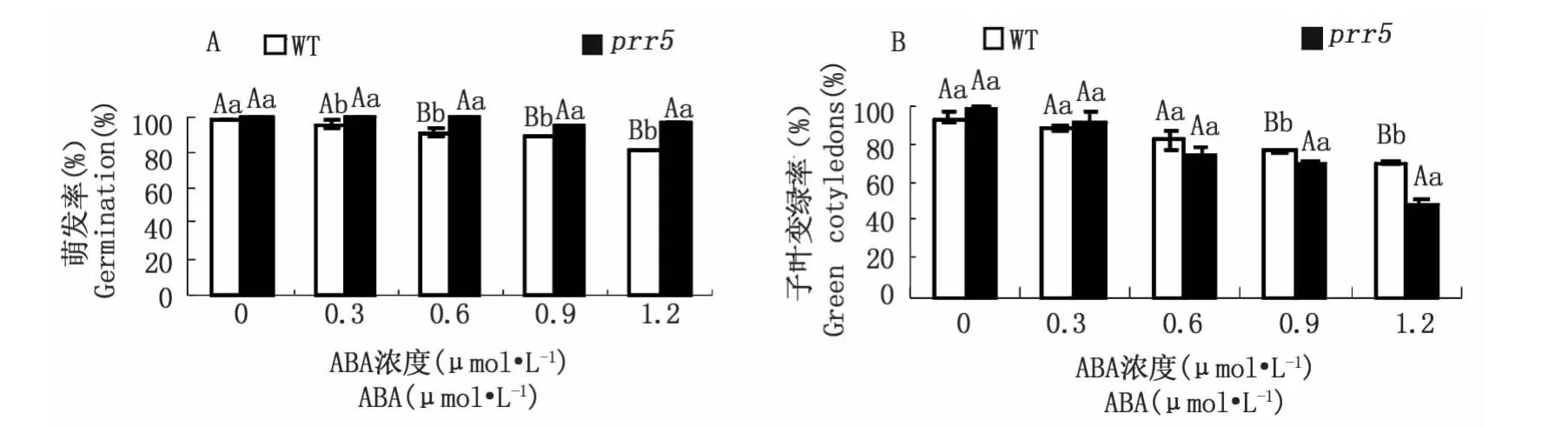
圖1 外源ABA對prr5突變體種子萌發(fā)率(A)及萌發(fā)后生長(子葉變綠率B)的影響Fig.1 Effects of exogenous ABA on germination inhibition(A)and post-germination growth(green cotyledons B)of prr5 mutant and wild-type seeds
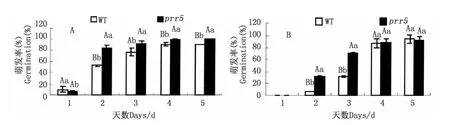
圖2 prr5突變體和野生型在ABA濃度為0.3(A)、0.9(B)μmol·L-1的 MS培養(yǎng)基上生長1、2、3、4、5 d 的萌發(fā)率Fig.2 Germination efficiency of wild type and prr5 mutant seeds on day 1 to day 5 with 0.3(A)and 0.9(B)μmol·L-1ABA treatment
2.2 ABA對prr5突變體幼苗主根伸長的影響
ABA對主根伸長有抑制作用,為了進一步從表型上探討PRR5基因是否與ABA信號轉(zhuǎn)導(dǎo)途徑關(guān)聯(lián),將在MS固體培養(yǎng)基上生長6 d的幼苗轉(zhuǎn)移到含不同濃度(0、0.1、1、10、50μmol·L-1)ABA 的 MS 固體培養(yǎng)基上,6 d后測量主根長度,從圖3可以看出,經(jīng)不同濃度ABA處理后,prr5突變體的主根長度都較野生型長,其中猶以1、50μmol·L-1ABA 濃度處理后突變體主根呈顯著、極顯著增長,即prr5也表現(xiàn)出對ABA弱敏感。
2.3 ABA對PRR5基因相對表達的影響
為了進一步驗證PRR5基因是否與ABA信號途徑關(guān)聯(lián),進一步從表達水平上檢測prr5是否受ABA調(diào)控。將擬南芥野生型7 d齡幼苗用不同濃度(0、1、10、50、100μmol·L-1)ABA 分別處理后,用實時定量PCR的方法對處理后幼苗中prr5的表達進行了分析,從圖4可以看出,低濃度ABA對PRR5基因表達的抑制作用明顯,隨濃度增加,抑制作用下降,但表達量均比未經(jīng)過ABA處理的要低,到100μmol·L-1濃度時跟未經(jīng)過處理的對照表達量基本一致,即PRR5基因表現(xiàn)出在高濃度時對ABA弱敏感。
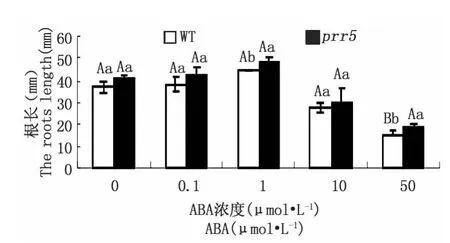
圖3 外源ABA對prr5突變體和野生型幼苗主根伸長的影響。野生型與prr5突變體幼苗在不含ABA的MS固體培養(yǎng)基上生長6天后轉(zhuǎn)移到含不同濃度(0,0.1,1,10,50μmol·L-1)ABA 的 MS 固體培養(yǎng)基上生長6天的根長生長情況Fig.3 Effects of exogenous ABA on the roots growth of prr5 mutant and wild-type seedlings.Sensitivity of root growth to ABA of the wild type and prr5 mutant,which are grown for 6 days on ABA-free medium and then incubated vertically for 6 days on medium with 0,0.1,1,10 or 50μmol·L-1ABA,respectively
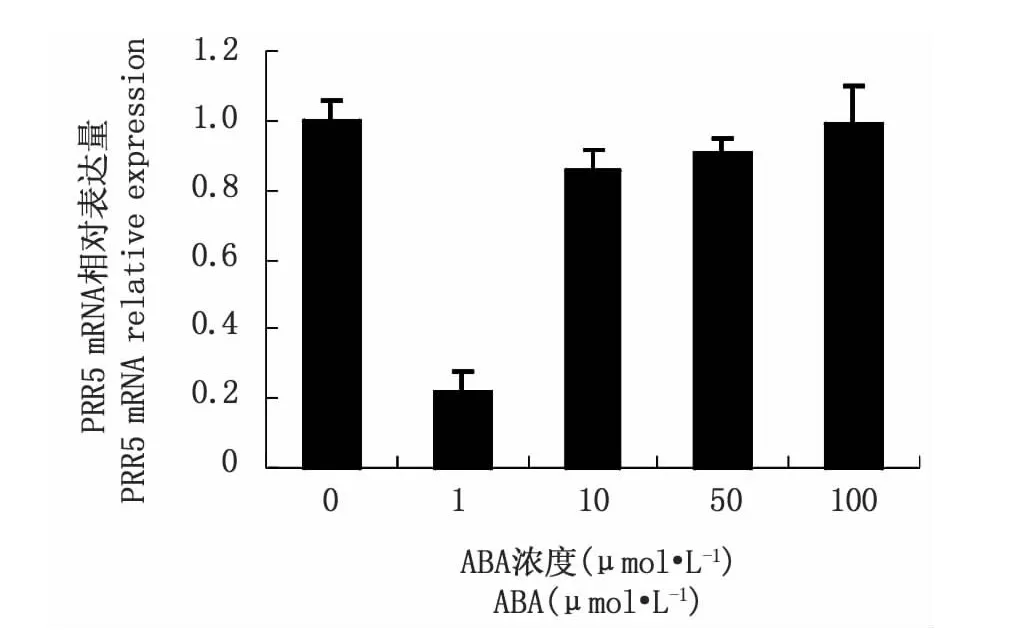
圖4 不同濃度ABA處理擬南芥幼苗后PRR5基因的表達Fig.4 The expression profiles of prr5 gene in seedling in response to different concentrations of ABA
2.4 NaCl處理對prr5突變體種子萌發(fā)的影響
脫落酸是一種重要的抗逆誘導(dǎo)因子,介導(dǎo)植物對逆境如干旱、鹽堿、低溫等生理反應(yīng)。為了研究prr5突變體對鹽脅迫是否產(chǎn)生反應(yīng),將突變體及野生型種子培養(yǎng)在含不同濃度(0、50、100、150 mmol·L-1)NaCl的MS固體培養(yǎng)基上,培養(yǎng)生長7 d后統(tǒng)計分析種子萌發(fā)率及子葉變綠率。圖5顯示,在不含NaCl的MS固體培養(yǎng)基上生長7 d后的種子萌發(fā)率、子葉變綠率與野生型相一致,但prr5突變體在含50、100、150 mmol·L-1NaCl MS固體培養(yǎng)基上的萌發(fā)率、子葉變綠率都比野生型的高,且均達到極顯著水平,猶以在含150 mmol·L-1NaCl培養(yǎng)基上生長變化最明顯。
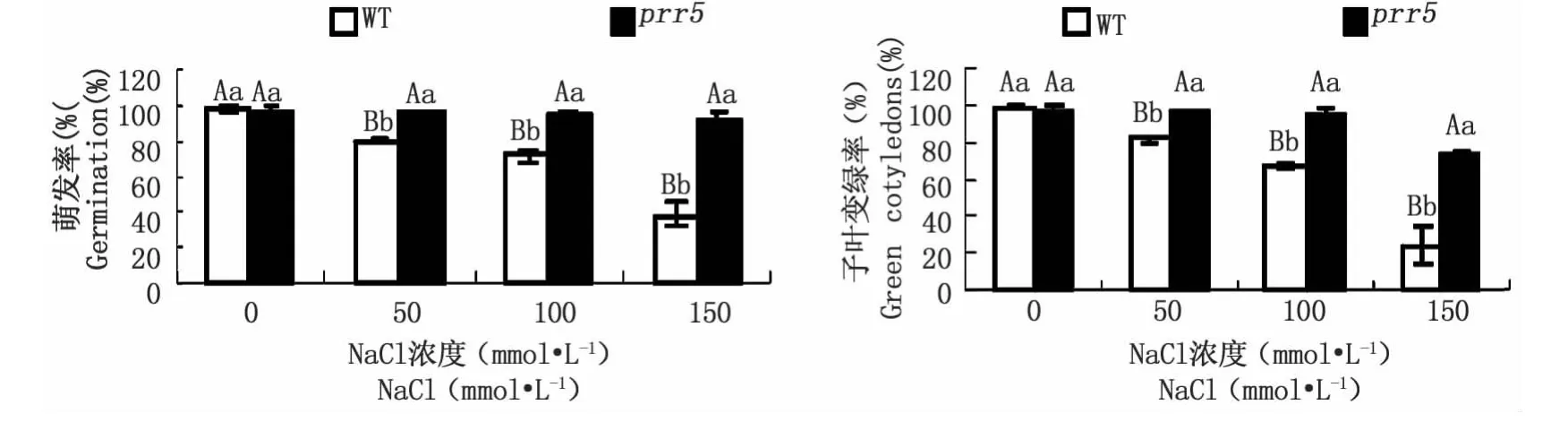
圖5 外源NaCl對prr5突變體和野生型萌發(fā)率(A)及萌發(fā)后生長(子葉變綠率B)的影響Fig.5 Effects of exogenous NaCl on germination(A)and post-germination growth(green cotyledons B)of prr5 mutant and wild-type seeds
3 討論
脫落酸是一種重要的植物激素,具有廣泛的生理功能,其信號轉(zhuǎn)導(dǎo)途徑是一個復(fù)雜的信號轉(zhuǎn)導(dǎo)網(wǎng)絡(luò)[12]。該信號網(wǎng)絡(luò)能被生物鐘節(jié)律調(diào)節(jié)。有研究發(fā)現(xiàn)生物鐘核心振蕩器成員PRR5、PRR7、PRR9的三缺失突變體prr9/7/5對ABA生物合成具有抑制作用。本研究用ABA處理prr5突變體及野生型,發(fā)現(xiàn)prr5突變體種子萌發(fā)率比野生型顯著或極顯著增高,主根比野生型長,即對ABA弱敏感。對野生型進行不同濃度ABA處理,發(fā)現(xiàn)PRR5基因表達受抑制,因此推測該基因可能為ABA信號轉(zhuǎn)導(dǎo)途徑的相關(guān)基因。同時,對prr5突變體與野生型種子進行NaCl處理,prr5突變體種子的萌發(fā)率比野生型極顯著增高,說明PRR5基因可能參與鹽脅迫調(diào)節(jié),由于ABA作為一種重要的抗逆誘導(dǎo)因子,介導(dǎo)植物對逆境如干旱、鹽堿、低溫等生理反應(yīng),NaCl處理實驗從側(cè)面驗證了PRR5基因?qū)BA的響應(yīng)。
TOC1 也屬于 PRRs基因家族[13,14]。有研究指出,TOC1是一個聯(lián)系生物鐘和干旱脅迫的分子開關(guān),TOC1-ox受干旱存活率比野生型低,TOC1 RNAi存活率比野生型的高[15]。Castells等[16]研究認(rèn)為TOC1調(diào)控ABA相關(guān)基因ABAR/CHLH/GUN5的表達是通過結(jié)合它的啟動子來進行的,同時發(fā)現(xiàn)toc1-2突變體經(jīng)ABA處理后的萌發(fā)率比野生型高,TOC1-ox突變體的萌發(fā)率比野生型低。因此,PRRs基因家族的其它成員有可能為ABA信號轉(zhuǎn)導(dǎo)途徑的相關(guān)基因。本試驗證明了PRR5參與了ABA信號轉(zhuǎn)導(dǎo)。prr5缺失突變體受ABA處理后的萌發(fā)率比野生型高,prr5缺失突變體的根長伸長也比野生型長,都表現(xiàn)出對ABA弱敏感,與上述toc1-2突變體對ABA處理后的結(jié)果相一致。因此,我們推測PRR5基因也可能通過調(diào)控ABA信號相關(guān)基因參與ABA信號途徑,我們將在今后作更深入的研究。
[1]KUCERA B,COHN M A,LEUBNER-METZGER G.Plant hormone interactions during seed dormancy release and germination[J].Seed Science Research,2005,15:281-307.
[2]KERMODE A R.Role of abscisic acid in seed dormancy[J].J Plant Growth Regulation,2005,224:319-344.
[3]FUKUSHIMA A,KUSANO M,NAKAMICHI N,et al.Impact of clock associated Arabidopsis pseudo-response regulators in metabolic coordination[J].Proceedings of the National Academy of Sciences,2009,106(17):7251-7256.
[4]CTAKAT A N,SAITO S,SAITO C T,et al.Phylogenetic footprint of the plant cock system in angiosperms:evolutionary processes of pseudo-response regulators[J].BMC Evolutionary Biology,2010,10:126.
[5]NAKAMICHI N,KIBA T,HENRIQUES R,et al.PSEUDORESPONSE REGULATORS 9,7,and 5 are transcriptional repressors in the Arabidopsis circadian clock[J].Plant Cell,2010,22:594-605.
[6]ROBERTSON F C,SKEFFINGTON A W,GARDNER M J,et al.Interactions between circadian and hormonal signaling in plants[J].Plant Molecular Biology,2009,69:419-427.
[7]王會,金宇騑,盧曼曼,等.擬南芥RopGEFs家族基因在外源脫落酸處理下的表達分析[J].植物生理通訊,2010,46(8):817-823.WANG Hui,JIN Yufei,LU Manman,et al.Expression analysis of ROPGEFS genes in Arabidopsis thaliana treated with ABA[J].PlantPhysiologyCommunications,2010,46(8):817-823.
[8]GUO X H,DENG K Q,WANG J,et al.Mutational analysis of Arabidopsis PP2CA2 involved in abscisic acid signal transduction[J].Molecular Biology Reports,2010,37:763-769.
[9]SAEZ A,ROBERT N,MOHAMMAD H,et al.Enhancement of abscisic acid sensitivity and reduction of water consumption in Arabidopsis by combined inactivation of the protein phosphatases type 2C ABI1 and HAB1[J].Plant Physiology,2006,114(4):1389-1399.
[10]劉實,趙小英,唐冬英,等.擬南芥CIPK1基因的功能初步分析[J].西北植物報,2007,27(6):1091-1095.LIU Shi,ZHAO Xiaoying,TANG Dongying,et al.The function of CIPK1 gene in Arabidopsis[J].Acta Botanica Boreal-Occidentalia Sinica,2007,27(6):1091-1095.
[11]鄧克勤,郭新紅,汪啟明,等.擬南芥磷酸酶基因亞細(xì)胞定位與組織表達[J].西北植物學(xué)報,2009,29(2):234-239.DENG Keqin,GUO Xinhong,WANG Qiming,et al.Subcellular localization and tissue expression pattern of Arabidopsis phosphatase gene[J].Acta Botanica Boreal-Occidentalia Sinica,2009,29(2):234-239.
[12]郭振清,郭曉強.脫落酸的信號轉(zhuǎn)導(dǎo)途徑[J].生命的化學(xué),2007,27(6):482-484.GUO Zhengqing,GUO Xiaoqiang.ABA signal transduction pathway[J].Chemistry of Life,2007,27(6):482-484.
[13]PARA A,F(xiàn)ARRE E M,IMAIZUMI T,et al.PRR3 is a vascular regulator of TOC1 stability in the Arabidopsis circadian clock[J].Plant Cell,2007,19:3462-3473.
[14]ERIKSSON M E,HANANO S,SOUTHERN M M,et al.Response regulator homologues have complementary light-dependent functions in the Arabidopsis circadian clock[J].Planta,2003,218:159-162.
[15]LEGNAIOLI T,CUEVAS J,MAS P.TOC1 functions as a molecular switch connecting the circadian clock with plant responses to drought[J].The EMBO Journal,2009,28:3745-3757.
[16]CASTELLS E,PORTOLéS S,HUANG W,et al.A functional connection between the clock component TOC1 and abscisic acid signaling pathways[J].Plant Signaling & Behavior,2010,5(4):409-411.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
