沙眼衣原體持續感染對Hela細胞TLR4/IL-6/STAT3信號通路的影響*
2013-11-10 11:14:38張寧潔王芙艷
激光生物學報 2013年6期
李 偉,林 琳,張寧潔,陳 恩,王芙艷,霍 治,余 平
(中南大學基礎醫學院免疫學系,湖南 長沙 410008)
沙眼衣原體是一類專性細胞內寄生的原核微生物,為臨床上導致失明的主要病因之一,也是最常見的性傳播性疾病(Sex transmitted disease,STD)病原體,WHO報道全世界每年有約9千萬新發STD病例是因感染Ct所致[1]。雖然Ct感染可用抗生素治療,但由于感染后癥狀常不明顯,甚至沒有自覺癥狀,往往會延誤診治,造成Ct持續感染[2],引起尿道炎、盆腔炎、輸卵管性不孕等多種疾病,甚至與HIV感染及腫瘤也有密切關系[3-5]。接種疫苗將是預防和控制Ct感染最有效的方式,但由于對機體的抗Ct感染免疫機制了解不夠,目前尚無有效的疫苗。
Toll-Like受體(Toll-like receptors,TLRs)主要表達在具有免疫功能的組織[6],例如:脾臟和外周血白細胞以及與外環境相通的呼吸和消化道、生殖道等,在早期固有免疫中對入侵病原微生物的識別發揮重要作用。這些進化保守的受體可識別表達在病原微生物上的高度保守的病原相關的分子模式(Pathogen-associated molecular pattern,PAMP),如酵母細胞壁上的甘露糖,各種細菌的脂多糖、多肽糖、胞壁酸等細胞壁成分,及鞭毛蛋白、細菌DNA和病毒的雙鏈RNA。TLRs受PAMPs刺激而啟動包括一些蛋白(例如MyD88和IRAK)的下游信號級聯,導致轉錄因子NF-kB的激活,誘導促炎/抑炎細胞因子和直接參與適應性免疫應答的效應細胞因子的分泌。
TLRs在Ct感染中的作用已被深入研究,主要集中于上皮細胞、樹突狀細胞(DC)及巨噬細胞(Mφ)上的 TLRs,尤其以 TLR2、TLR4的作用最為突出[7,8]。如 Ct的 LPS 及 cHSP60 可經 TLR2 或 TLR4途徑被上皮細胞、樹突狀細胞等識別,誘生IL-6、IFN-β等,進而對獲得性免疫進行調節[9]。目前已有的關于衣原體感染后TLRs表達變化的研究證據在不同的研究報道中并不一致,其原因可能與各研究中使用的衣原體菌株及感染靶細胞株不同有關,也有可能與所建立的感染模型不同(急性/持續性)有關,但目前尚無這方面的研究報道。
已有研究證實在Ct感染后,Ct的某些組分可經TLRs信號通路刺激上皮細胞、抗原提呈細胞(APC)等產生前炎癥因子,而我們認為在感染進程的不同階段,TLRs在靶細胞上的表達可能隨著感染的進程變化而變化,進而影響前炎癥因子的產生,最終在免疫清除及慢性炎癥中表現出不同的生物學功能。我們利用Hela細胞分別建立Ct急性感染及持續性感染模型,通過熒光定量RT-PCR、ELISA等研究策略比較Ct不同感染狀態下TLR4、IL-6以及其下游STAT3表達量的變化,初步探索TLRs參與衣原體持續感染的可能機制,為Ct感染及炎癥的控制提供新的實驗依據。
1 材料與方法
1.1 實驗材料及試劑
人宮頸癌上皮細胞株Hela及Ct L2血清型為本室保存;IFN-γ購自 SIGMA;DMEM、小牛血清購自Hyclone;DEPC、Trizol、異丙醇購自北京鼎國;MMLV逆轉錄試劑盒購自Fermantas;qRT-PCR試劑購自BioRad;IL-6 ELISA試劑盒購自R&D。
1.2 細胞培養和Ct急性感染模式及持續性感染模式的建立
(1)Hela細胞用含有10%小牛血清的DMEM在37℃,5%CO2的培養箱中培養。參照文獻[9]進行Ct感染、增殖和滴定,分裝的Ct保存在-80℃。
(2)建立急性感染模式:計數2×105個Hela細胞接種于60 mm細胞培養皿,24 h后感染Ct L2血清型 (MOI=5,MOI multiplicity of infection,感染復數)。感染細胞在37℃、5%CO2培養箱中培養,分別在感染后不同時間點收取標本。
(3)建立持續性感染模式:計數2×105個Hela細胞接種于60 mm細胞培養皿,接種24 h后感染Ct L2血清型 (MOI=5),參照前期實驗結果,在2 h后加入100 U/mL的IFN-γ。感染細胞在37℃5%CO2培養箱中培養,分別在感染后不同時間點收取標本。
1.3 熒光定量 PCR技術檢測 TLR4、IL-6以及STAT3轉錄水平
分別在Ct急性感染及持續性感染Hela細胞6 h、18 h、24 h和48 h(分別代表Ct發育周期的極早期、早期、中期和晚期)后收集細胞樣本,同時收集相同時間點未感染細胞樣本作為對照,參照使用說明書用TRIzol法抽提總RNA。將RNA樣本用MMLV試劑盒逆轉錄合成cDNA,保存在-80℃。
并利用Primer2.0及Primer-Blast設計待測基因的引物(表1)用于qRT-PCR檢測mRNA表達量。所采用的qRT-PCR體系(20μL)為:

qRT-PCR循環條件如下:95℃變性2 min,后以94℃變性15 s,55℃退火15 s,40個循環,每一循環結束后記錄熒光值,擴增完畢后繪制熔解曲線。采用△△Ct法計算各目的基因 mRNA的表達變化。使用BioRad CFX manager 2.0分析不同感染狀態下基因的差異表達情況。
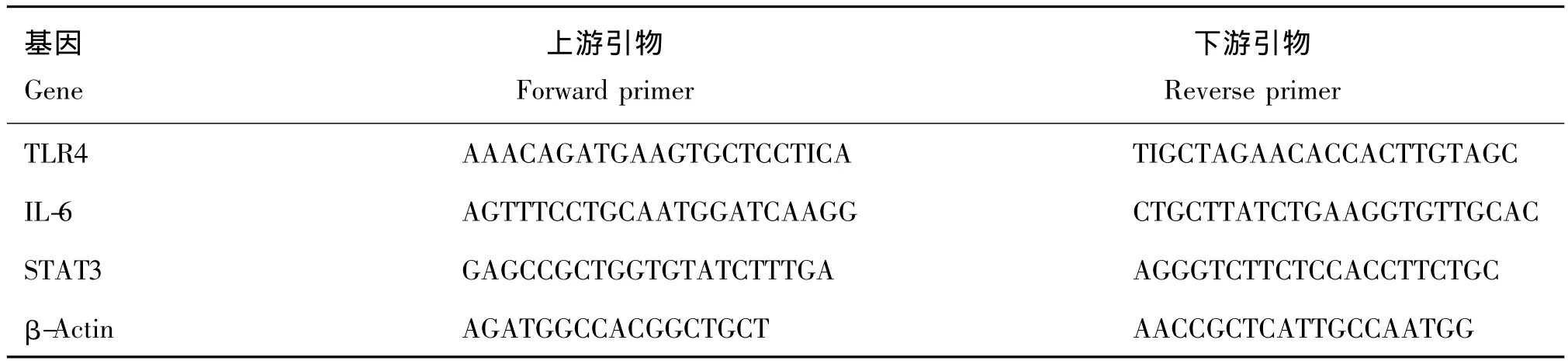
表1 qRT-PCR引物序列Tab.1 The primers for qRT-PCR
1.4 ELISA檢測IL-6分泌水平
分別在Ct急性感染及持續性感染Hela細胞6 h、12 h、24 h、36 h和48 h后收集培養上清,同時收集相同時間點未感染培養上清樣本作為對照,分裝后保存在-80℃。用雙抗夾心ELISA方法檢測上清液中IL-6濃度。操作根據試劑盒說明書進行。
1.5 統計分析
所有資料處理均使用SPSS13.0軟件進行分析,檢測結果以表示,組間比較采用獨立樣本的t檢驗,以P<0.05為差異有統計學意義。
2 實驗結果
2.1 成功構建Ct持續性感染模型
在前期實驗基礎上,我們利用IFN-γ建立了Ct L2持續感染Hela細胞的模型(圖1)。
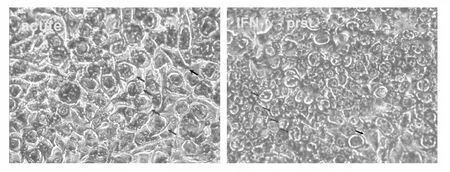
圖1 光學顯微鏡下觀察L2急性感染(acute)及IFN-γ誘導的持續性感染(IFN-γ-prst)的Hela細胞(姬姆薩染色,40×10)Fig.1 Observed under optical microscope:acute infection(acute)and IFN-γ-induced persistent infection(IFN-γ-prst)of L2 in Hela cells(Giemsa staining,40 × 10)
2.2 不同感染組中 TLR4、IL-6和STAT3的表達變化
利用qRT-PCR方法發現,L2感染后Hela細胞的TLR4轉錄水平出現上調,且隨著感染時間延長,上調程度增加;與急性感染相比較,IFN-γ誘導的L2持續感染狀態下Hela細胞的 TLR4轉錄水平上調改變得更為顯著(圖2)。而IL-6轉錄水平的表達也表現出相似的改變,在持續性感染狀態下Hela細胞內IL-6轉錄水平比未感染組、急性感染組顯著增加,并呈時間依賴性增加,而且與TLR4的轉錄表達上調趨勢相一致(圖3)。STAT3的轉錄水平在持續性感染18 h明顯增強,之后隨時間推移轉錄水平逐漸下調,但總體轉錄水平比未感染組和急性感染組顯著增高(圖 4)。
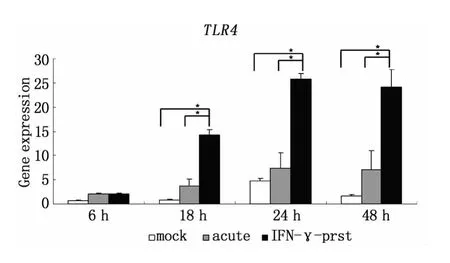
圖2 qRT-PCR技術檢測L2不同感染模式下,Hela細胞不同時間點TLR4的轉錄水平mock:未感染;acute:急性感染模型;IFN-γ-prst:IFN-γ誘導的持續感染模型 (*P<0.05)Fig.2 qRT-PCR detection of TLR4 transcription level in Hela cells in different pattern of L2 infection at different time points(*P <0.05)mock:mock infecion;acute:acute infection;IFN-γprst:IFN-γ-induced persistent infection
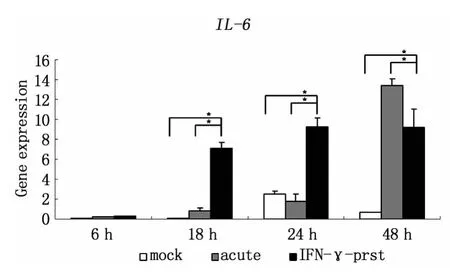
圖3 qRT-PCR技術檢測L2不同感染模式下,Hela細胞不同時間點IL-6的轉錄水平mock:未感染;acute:急性感染模型;IFN-γ-prst:IFN-γ誘導的持續感染模型 (*P<0.05)Fig.3 qRT-PCR detection of IL-6 transcription level in Hela cells in different pattern of L2 infection at different time points(*P <0.05)mock:mock infection acute;acute infection:IFN-γprst:IFN-γ-induced persistent infection
2.3 不同感染組中IL-6分泌水平
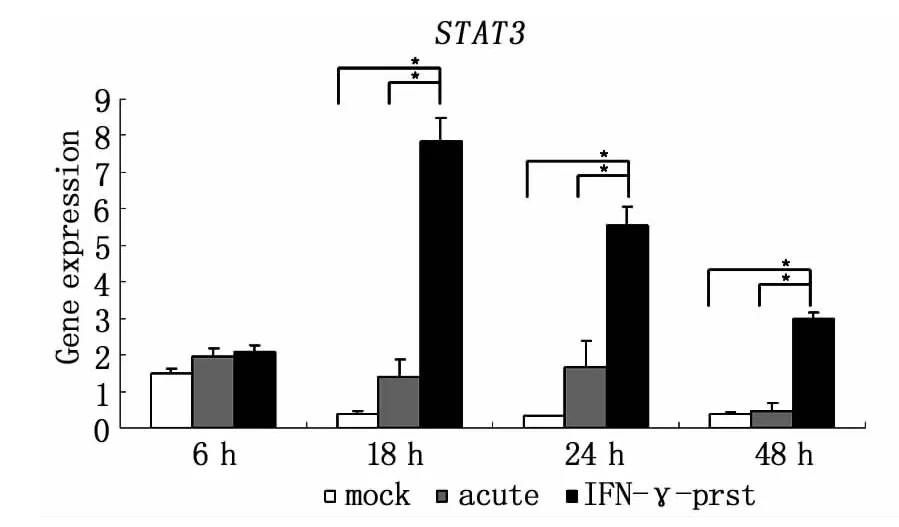
圖4 qRT-PCR技術檢測L2不同感染模式下,Hela細胞不同時間點STAT3的轉錄水平mock:未感染;acute:急性感染模型;IFN-γ-prst:IFN-γ誘導的持續感染模型(*P<0.05)Fig.4 qRT-PCR detection of STAT3 transcription level in Hela cells in different pattern of L2 infection at different time points(*P <0.05)mock:mock infection;acute:acute infection;IFN-γprst:IFN-γ-induced persistent infection
利用ELISA方法檢測細胞因子分泌發現,L2感染后Hela細胞的IL-6分泌水平出現明顯上調,且隨著感染時間延長,上調程度增加;與急性感染相比較,IFN-γ誘導的L2持續感染狀態下Hela細胞的IL-6分泌水平上調改變得更為顯著(圖4)。

圖5 ELISA檢測L2不同感染模式下,培養Hela細胞不同時間后上清中IL-6的濃度mock:未感染;acute:急性感染模型;IFN-γ-prst:IFN-γ誘導的持續感染模型(*P<0.05)Fig.5 ELISA detection of IL-6 secretion level in Hela cells supernatant in different pattern of L2 infection at different time points(*P <0.05)mock:mock infection;acute:acute infection;IFN-γprst:IFN-γ-induced persistent infection
3 討論
Ct在宿主細胞內的繁殖呈特殊的二相發育周期,分別為有感染性、無分裂能力的原體(Elementary body,EB)及快速生長但無感染性的網狀體(Reticulate body,RB)。RB以二分裂方式增殖后再度分化成EB,EB從細胞釋放后再次感染新的宿主細胞,整個發育周期約需 48-72 h[2]。在饑餓、抗生素、IFN-γ作用以及抗感染免疫等條件下,Ct在宿主細胞內會形成非典型感染狀態,電鏡下表現為既不同于原體也不同于始體的細胞,被稱為異常RB,為目前公認的Ct持續感染顯微結構[10]。在前期實驗基礎上,我們利用IFN-γ建立了Ct L2持續感染Hela細胞的模型。光鏡下觀察發現持續感染狀態下的L2包涵體體積小于急性感染狀態下的L2包涵體,但更細微的亞細胞結構則需要在電鏡下觀察。
TLRs是廣泛存在于固有免疫細胞上的一類模式識別受體(Pattern-recognition receptors,PRRs),可識別多種PAMPs[6]。在病原體入侵的幾分鐘內,各種TLRs即可通過相同或不同的胞內信號轉導途徑誘導一系列基因活化,致使細胞分泌炎癥細胞因子和趨化因子,誘發炎癥反應,產生固有免疫應答,并在適應性免疫應答中起重要的調節作用。已有研究證實,急性感染狀態下,TLR4信號途徑可識別Ct來源的 cHSP60、LPS 等 PAMPs,誘生 IFN-γ、IL-6、IL-10 等多種炎癥因子,在調控固有免疫細胞的活性、控制或消除感染中起重要作用[7,8]。我們研究發現,隨著Ct感染時間延長,Hela細胞的TLR4轉錄水平明顯升高,與Ct急性感染相比較,持續感染模型中的變化更為顯著,提示Ct持續感染狀態下宿主細胞內的TLR4信號通路受到了調控,但其具體調控機制及作用尚不明確。
現已知TLRs途徑可誘導細胞因子產生。TLR4單抗可有效阻斷肺炎衣原體HSP10作用的人單核-巨噬樣細胞 THP-1細胞分泌 IL-6[11]。還有研究證實,TLRs信號與 IL-6間存在負反饋環[12]。我們研究發現,Ct不同感染模式下,Hela細胞的TLR4轉錄水平隨時間變化趨勢與IL-6的分泌水平一致,但兩者之間是單純的因果關系還是存在更為復雜的負反饋調控機制,還需要進一步的實驗分析。
Ct感染的病理特點是炎癥反應,隨著反復持續感染,炎癥加重,最終導致組織損傷和瘢痕形成。Ct感染的主要靶細胞是泌尿生殖道上皮細胞和眼瞼粘膜上皮細胞,它們可以產生大量炎癥因子,尤其是IL-1、IL-8 和 IL-6 等[7-9],炎癥因子一方面可以直接介導宿主的固有免疫應答,參與宿主抵抗Ct感染的第一道防線;另一方面可以導致組織細胞的病理損傷。因此,在感染過程中Ct與這些炎癥因子的相互作用關系是疾病進展的一個重要環節,研究這些炎癥因子產生的機制有助于理解Ct的致病機制,對疾病防治也有重要意義。IL-6作為具有最廣泛的生物學效應的細胞因子之一,同時具有促炎和抗炎的雙向作用。在內皮細胞中,IFN-γ介導的SOCS3表達導致了IL-6誘導的STAT3活化受抑,進而降低了IL-6依賴的抗炎基因表達,使得IL-6的生物學作用由抗炎向促炎發生轉變,參與了內皮細胞炎癥損傷[13,14]。而關于IL-6在Ct感染中的促炎和抗炎作用目前并無統一觀點,之前的研究發現在衣原體的持續感染狀態下有持續的IL-6的分泌[15],我們的研究也證實在Ct感染后,尤其是持續感染狀態下,宿主細胞持續高表達IL-6,且隨著感染時間延長IL-6分泌量增加,提示IL-6的產生可能促進了細菌生長。但是Ct誘導IL-6的產生機制以及IL-6在Ct感染中的具體作用還不清楚,在Ct反復持續感染宿主過程中,IL-6是發揮促炎作用加重炎癥還是發揮抑炎作用參與免疫調節,以及是否有助于Ct逃避免疫清除,目前仍不清楚。
STAT3是信號傳導和轉錄活化因子(Signal transducer and activator of transcription,STAT)家族的成員之一,IL-6的信號轉導主要依賴于STAT3的參與[16]。IL-6與其膜受體結合后,激活STAT3,使其二聚化進入核內啟動多種基因轉錄活化,啟動多種效應,包括細胞增殖反應、急性炎癥反應以及細胞凋亡等[17]。本研究發現在用 IFN-γ建立的 Ct持續感染的模型中,與Ct急性感染相比,Hela細胞的STAT3轉錄水平顯著升高,且呈現出在感染早期(18 h之內)急劇升高,18 h后逐漸下調的變化趨勢;有趣的是,這一變化趨勢與IL-6的mRNA及蛋白質表達變化并不完全一致,推測可能是某些目前尚不明確的機制(如STAT1的拮抗作用)調控了STAT3的轉錄及轉錄后的蛋白活性。但是有關Ct持續性感染后的STAT3下游信號尚不清楚,STAT3信號參與促炎還是抑炎,介導細胞損傷還是組織保護,因此我們的工作初步探索了TLRs可能參與衣原體持續感染后IL-6/STAT3信號通路的調控,更具體的分子機制還需要我們進一步的研究。
參考文獻
[1]AGRAWAL T,VATS V,SALHAN S,et al.The mucosal immune response to Chlamydia trachomatis infection of the reproductive tract in women[J].J Reprod Immunol,2009,83(1-2):173-178.
[2]HAMMERSCHLAG M R.The intracellular life of chlamydiae[J].Semin Pediatr Infect Dis,2002,13(4):239-248.
[3]SHAW J L,HORNE A W.The paracrinology of tubal ectopic pregnancy[J].Mol Cell Endocrinol,2011,358(2):216-222.
[4]GHOSH A,DHAWAN B,CHAUDHRY R,et al.Genital mycoplasma and Chlamydia trachomatis infections in treatment naive HIV-1 infected adults[J].Indian J Med Res,2011,134(6):960-966.
[5]DELUCA G D,BASILETTI J,SCHELOVER E,et al.Chlamydia trachomatis as a probable cofactor in human papillomavirus infection in aboriginal women from northeastern Argentina[J].Braz J Infect Dis,2011,15(6):567-572.
[6]LAVELLE E C,MURPHY C,O'NEILL L A,et al.The role of TLRs,NLRs,and RLRs in mucosal innate immunity and homeostasis[J].Mucosal Immunol,2010,3(1):17-28.
[7]CAPPELLO F,CONWAY DE MACARIO E,et al.Chlamydia trachomatis infection and anti-Hsp60 immunity:the two sides of the coin[J].PLoS Pathog,2009,5(8):e1000552.
[8]AGRAWAL T,BHENGRAJ A R,VATS V,et al.Expression of TLR 2,TLR 4 and iNOS in cervical monocytes of Chlamydia trachomatis-infected women and their role in host immune response[J].Am J Reprod Immunol,2009,66(6):534-543.
[9]SRIVASTAVA P,JHA R,BAS S,et al.In infertile women,cells from Chlamydia trachomatis infected sites release higher levels of interferon-gamma,interleukin-10 and tumor necrosis factor-alpha upon heat-shock-protein stimulation than fertile women[J].Reprod Biol Endocrinol,2008,6:20.
[10]SKILTON R J,CUTCLIFFEN L T,BARLOW D,et al.Penicillin induced persistence in Chlamydia trachomatis:high quality time lapse video analysis of the developmental cycle[J].PLoS One,2009,4(11):e7723.
[11]ZHOU Z,WU Y,CHEN L,et al.Heat shock protein 10 of Chlamydophila pneumoniae induces proinflammatory cytokines through Toll-like receptor(TLR)2 and TLR4 in human monocytes THP-1[J].In vitro Cell Dev Biol Anim,2011,47(8):541-549.
[12]YILMA A N,SINGH S R,FAIRLEY S J,et al.The anti-inflammatory cytokine,interleukin-10,inhibits inflammatory mediators in human epithelial cells and mouse macrophages exposed to live and UV-inactivated Chlamydia trachomatis[J].Mediators Inflamm,2012,520174.
[13]HU X,IVASHKIV L B.Cross-regulation of signaling pathways by interferon-gamma:implications for immune responses and autoimmune diseases[J].Immunity,2009,31(4):539-550.
[14]HONG F,JARUGA B,KIM W H,et al.Opposing roles of STAT1 and STAT3 in T cell-mediated hepatitis:regulation by SOCS[J].J Clin Invest,2002,110(10):1503-1513.
[15]MPIGA P,MANSOUR S,MORISSET R,et al.Sustained interleukin-6 and interleukin-8 expression following infection with Chlamydia trachomatis serovar L2 in a HeLa/THP-1 cell coculture model[J].Scand J Immunol,2006,63(3):199-207.
[16]GAO B,WANG H,LAFDIL F,et al.STAT proteins-key regulators of anti-viral responses,inflammation,and tumorigenesis in the liver[J].J Hepatol,2012,57(2):430-441.
[17]MOH A,IWAMOTO Y,CHAI G X,et al.Role of STAT3 in liver regeneration:survival,DNA synthesis,inflammatory reaction and liver mass recovery[J].Lab Invest,2007,87(10):1018-1028.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
光學精密工程(2016年6期)2016-11-07 09:07:19
學苑創造·A版(2015年11期)2016-01-14 09:03:27
核科學與工程(2015年4期)2015-09-26 11:59:03
中國火炬(2010年8期)2010-07-25 11:34:30
