原子力顯微鏡研究不同刺激條件下T細胞的形態和生物力學特性*
2013-11-10 11:14:32邢曉波朱德斌王海燕蔡繼業
激光生物學報 2013年6期
關鍵詞:結構
邢曉波,黃 訓,朱德斌,王海燕,蔡繼業
(1.暨南大學化學系,廣東廣州 510632;2.華南師范大學生物光子學研究院激光生命科學研究所教育部重點實驗室,廣東廣州 510631;3.暨南大學材料科學與工程系,廣東廣州 510632;4.廣州計量檢測技術研究院,廣東 廣州 510663)
T淋巴細胞是在胸腺中分化成熟的淋巴細胞,故稱胸腺依賴性淋巴細胞(Thymus-dependent lymphocyte),簡稱T細胞。T細胞是淋巴細胞中數量最多,功能最復雜的一類細胞,是承擔細胞免疫的活性細胞。T細胞具有抗原識別受體,因此它們必需經抗原刺激才能活化而發揮其效應細胞的作用,為特異性細胞免疫。大量的研究表明[1],病原體感染、腫瘤的發生、自身免疫性疾病的發生以及有關組織器官移植排斥等都與T細胞抗原識別和活化異常或偏離相關。抗原是一類能刺激機體產生抗體或致敏淋巴細胞,并能與相應的抗體或致敏淋巴細胞特異性結合,發生免疫應答的物質,是引起特異性免疫的激發原[2]。當T淋巴細胞識別出細胞表面存在抗原多肽時,免疫系統便將其認為異種細胞(如腫瘤細胞,病毒感染的細胞)并開始攻擊。因此,T細胞的抗原識別和活化可以直接影響整個免疫應答的性質、效能和結果,在人體免疫反應中具有核心作用。T淋巴細胞活化主要表現為細胞分裂增殖,克隆擴增,并出現分化,由靜止狀態轉變為效應細胞并執行各種功能[3]。而T淋巴細胞的靜息和(不同時間的)活化期,表達在細胞膜的膜表面抗原分子的種類和數量也是不同的,同時,T淋巴細胞的形態、超微結構和生物力學特性會發生一系列的變化[4-5]。由于細胞膜表面受體和表達蛋白等都是生物大分子,特征尺寸在10~100 nm量級,細胞力學特性的變化在皮牛頓量級,這就需要有高分辨率的納米手段和高靈敏度的測力手段對其進行研究。隨著生物納米技術的發展和掃描探針顯微術的發展,使得原子力顯微鏡(AFM)能夠分析細胞和生物材料的納米結構和生物力學特性[6-8]。
機體的器官、組織、細胞和生物大分子在一定范圍力學作用下發生相應的形態和功能改變,這是機體對力學刺激的響應過程,而機體對一定范圍力學刺激的自適應對于維持正常生理功能具有重要作用。研究力對細胞的作用,是揭示器官、組織生物力學特性的基礎,也是進一步研究細胞內生物大分子的生物力學特性的出發點。細胞粘附包括細胞之間的粘附以及細胞與基質之間的粘附,其中細胞之間粘附是細胞間信息交流的一種形式,細胞與基質的粘附與細胞的遷移和運動密切相關[9],其過程涉及復雜的細胞生物化學、機械性能、表面鍵合、電荷、自由能、疏水性等因素[10],細胞的彈性是細胞維持其形態和承受一定的壓力所必須的,因此對研究T細胞活化前后粘附力和彈性模量的變化,對于揭示其生物學功能具有重要的意義。大量研究表明,力可以通過影響細胞內基因表達和蛋白質的合成來調節細胞功能,在細胞的生理、病理過程中發揮著重要作用[11]。
本實驗基于AFM的力譜測量功能,對靜止(resting)T淋巴細胞以及經超抗原(SEA)或植物凝集素(PHA)活化后T淋巴細胞的形態結構、表面超微結構成像和探測研究,并通過比較不同狀態下T淋巴細胞表面粘附力和硬度的變化,探討T淋巴細胞形態變化與粘附行為和細胞硬度之間的關系,以便能更深入地了解T淋巴細胞形態結構和細胞行為之間的關系。
1 材料與方法
1.1 實驗材料及主要儀器
健康人外周血,收集于肝素鈉抗凝的試管內;超抗原(StaphylococcalenterotoxinA,SEA)、植物凝集素(Phytohaemagglutinin,PHA)、戊二醛均購自 Sigma公司;人外周血淋巴細胞分離液,人T細胞富集液(RosetteSep?HumanT Cell Enrichment Cocktail)均購自AXIS SHIELD PoC AS公司;RPMI-1640、胎牛血清(Fetal bovine serum,FBS)及β-巰基乙醇等細胞培養試劑均為GibcoBR公司產品;其它試劑均采用國產分析純,實驗用水為超純水。原子力顯微鏡 (Autoprobe CP research,thermomicroscopes公司,美國)。
1.2 T細胞的分離和活化
將2mL健康人外周血與肝素抗凝劑混勻以抗凝。具體分離步驟如下:1)將100μL T細胞富集液與2mL全血輕輕混勻,并在室溫下孵育20 min;2)再用等體積(2mL)的含2%牛血清白蛋白(BSA)的PBS液輕輕稀釋混勻;3)將稀釋后的血輕輕鋪在3mL的密度分離液(Ficoll)上面(注意讓其分層,而不能混合),然后室溫下以1200 g的速度離心20 min;4)取密度分離液與血漿之間的細胞富集層(白色),細胞用2%BSA+PBS溶液離心洗兩次(425 g,10 min)。將分離的細胞重懸于RPMI 1640培養 液 培 養(含 10%FBS 及 50μmol/L β-巰 基 乙醇),調整細胞密度為2×106cells/mL。臺盼藍染色觀察細胞活力,活細胞數占98%。T淋巴細胞設置3組:Resting組(不添加任何外源刺激劑)、SEA(10μg/mL)刺激組和 PHA(10μg/mL)刺激組。將細胞在24孔板接種后置于37℃、體積分數為5%的CO2培養箱中培養48 h。
1.3 樣品的制備
分別取Resting、SEA活化和PHA活化48 h的T淋巴細胞,滴于干凈的蓋玻片上,使其自然鋪展,吸附10 min,然后用1.5%的戊二醛固定10~15 min,用蒸餾水沖洗3次,室溫自然干燥。
1.4 AFM成像及力譜測量
將制備好的樣品置于AFM的XY掃描臺上,用監視器定位所要掃描的樣品區域,在空氣中室溫下,利用接觸模式成像。實驗采用100μm掃描器,UL20B硅探針,力常數為2.8 N/m。AFM圖像僅經過自帶軟件(IP 2.1版)的平滑處理,以消除掃描方向上的低頻背景噪音。
1.5 統計學分析
利用SPSS 13.0進行力曲線相關數據以及細胞的各項物理參數統計分析,采用 t檢驗方法,P<0.05被認為是有統計學意義的。取自10個細胞的所有數據用平均值±標準方差表示。
2 結果
2.1 倒置熒光顯微鏡觀察活化前后T細胞的形態
圖1a和1b為未加刺激劑培養了48 h時T細胞在倒置熒光顯微鏡下的形態,細胞呈規則的圓形,從40倍放大成像可以看出細胞與細胞之間沒有連接到一起(圖1b)。從圖1c和1d可以看出經PHA刺激48 h時,細胞發生凝集,出現細胞團如圖1c中紅色虛線所示,細胞團大小不一如圖1d中紅色虛線所示。從圖1e和1f也可以看到SEA活化后細胞也發生凝集但是與PHA活化相比形成的細胞團較小,與對照組相比細胞分布不均勻,有的地方細胞密度較大有的地方細胞密度較小。從40倍的放大圖(圖1f)可以看到有些細胞與細胞連接到一起。
2.2 T細胞形貌和膜納米結構的AFM表征
首先,研究了靜息的 T細胞的形貌結構與膜超微結構(如圖2)。從圖2a可以看出靜息的T細胞為規則的圓形。其中圖2b是圖2a的三維圖像,圖2c為誤差信號模式圖,在這種成像模式下許多特化結構如細胞邊緣的偽足等可以更加清楚的顯示出來。從其剖面圖(圖2d)可以看出靜息的T細胞高度為1.86μm,半高寬為 5.0μm。圖 2e 和圖 2f是膜表面納米結構圖像,其中圖2f是圖2e的三維圖。靜息的T細胞表面相對比較光滑(圖2e和2f)。為了在納米空間對靜息的和不同活化劑活化的T細胞的細胞膜化學成分的變化進行分析,我們利用AFM的橫向力顯微(Lateral force microscopy,LFM)功能,對細胞膜表面的變化進行了成像。根據材料表面物理特性如表面摩擦力的差異,AFM在橫向力成像模式(lateral force,LF)下可以獲得材料表面具有不同物理特性的結構分布[9]。圖2g為靜息的T細胞膜納米結構的LFM圖,圖中顏色由紅到藍表示摩擦力由大變小,從圖中可以看出細胞表面摩擦力的大小分布并不是非常均勻的,這主要是由于細胞表面顆粒的高度和細胞膜成分的差異引起的。圖2h是膜表面顆粒粒徑的分布圖,其表面顆粒粒徑分布在20~60 nm之間,其中主要集中在40 nm。
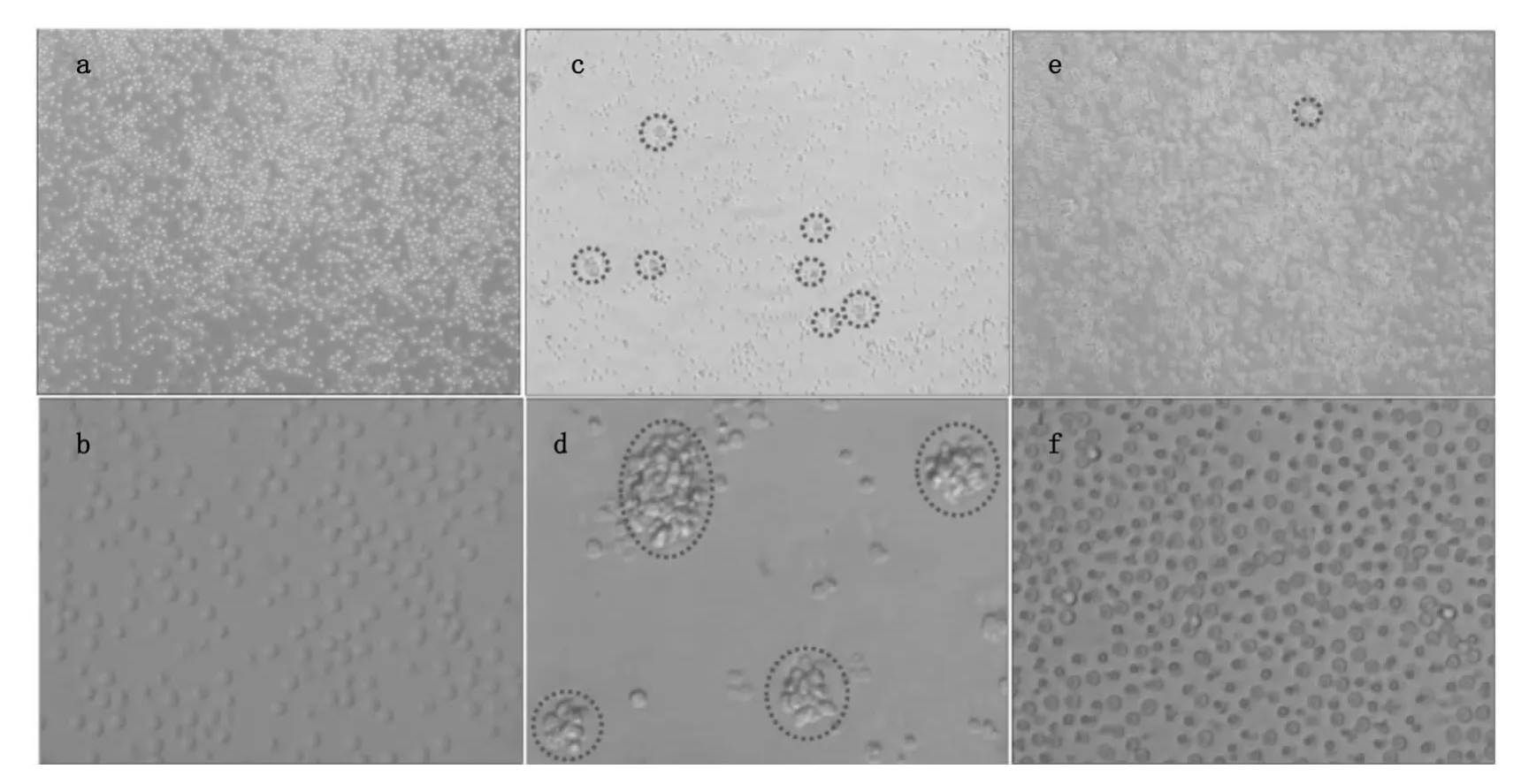
圖1 倒置熒光顯微鏡下T細胞活化前后的形態。圖(a)和圖(b)是沒有加刺激劑的T細胞,圖(c)和圖(d)是T細胞經PHA刺激48 h時的形態,圖(e)和圖(f)是T細胞經SEA刺激48 h時的形態Fig.1 Microscopic images of T cells before and after SEA and PHA treatment for 48 h.Concentrations of PHA and SEA used in this study were all 10μg/mL.(a,b,c,d,e and f)images of T cells before(a,b)and after PHA(c,d)and SEA(e,f)treatment

圖2 靜息的T細胞的形貌結構與膜納米結構。圖b是圖a的三維圖像,圖c為誤差信號模式圖,圖d是細胞a的剖面圖,圖e和圖f是膜表面超微結構圖像,圖g是橫向力(LFM)模式圖,圖k是膜表面顆粒粒徑的分布圖Fig.2 Representative AFM morphological images of resting T cells.(b)3-D image of(a).(c)The signal error mode image.(d)Height profile generated along the black line in(a).(e,f)Ultrastructural images of cell membrane.(g)Lateral force images of(e).(h)The diameter of particles distribution of membrane surface particle in(e)
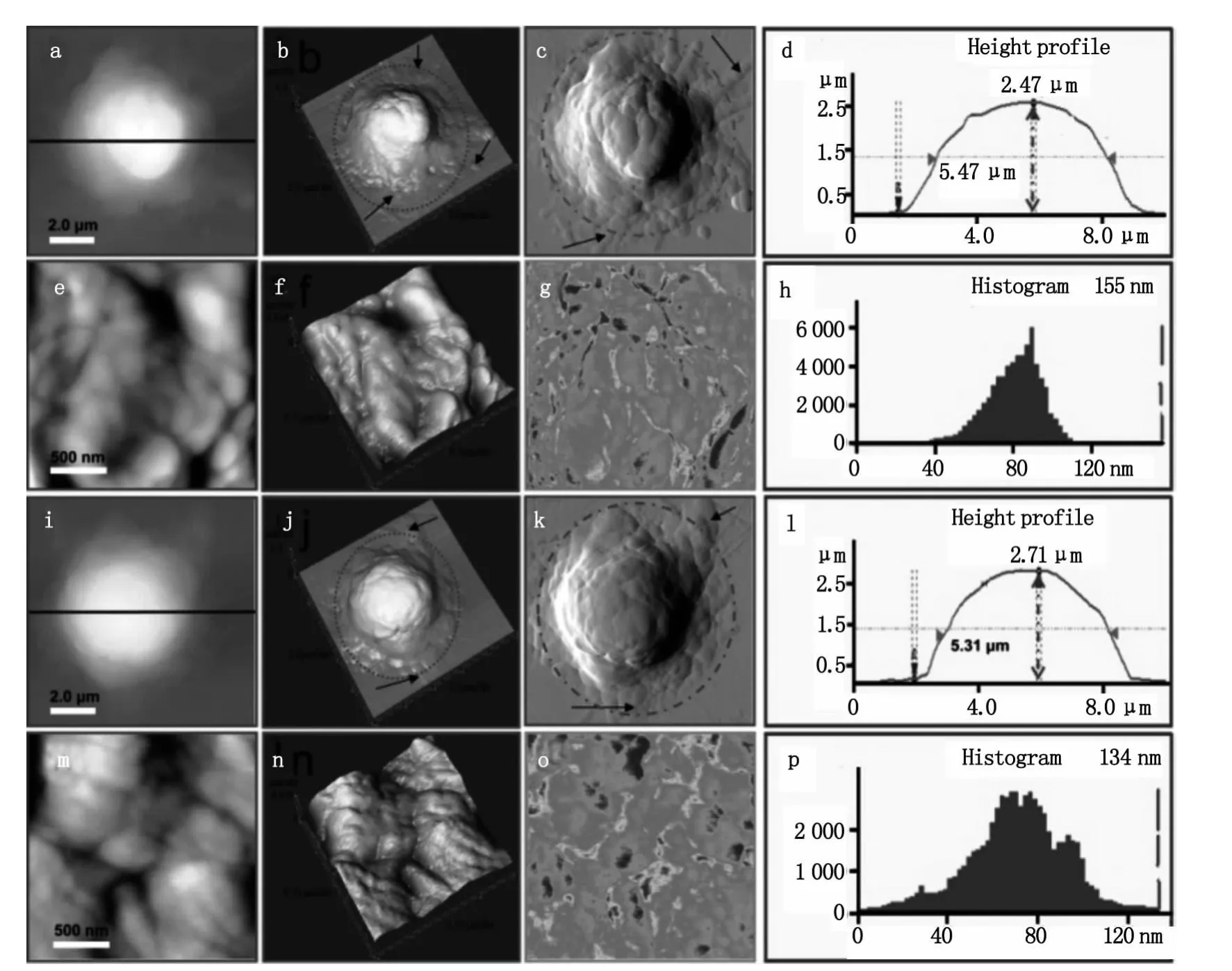
圖3 經刺激劑處理后的活化的T細胞的具有代表性的形貌圖和膜納米結構圖像。圖a-h顯示了PHA活化的T細胞的形貌和膜納米結構結構,圖i-p顯示了SEA活化的T細胞的形貌和膜納米結構Fig.3 Representative AFM images of activated T cells.(a-h)and(i-p)show the morphology and nanostructure of T cells which are activated by PHA and SEA,respectively
如圖3所示是用PHA和SEA分別刺激活化后的T細胞的AFM觀察結果。圖3a到3c是PHA活化后T細胞的形貌圖,從三維圖(圖3b)和誤差信號圖(圖3c)可以明顯的看到活化后細胞周圍的蹼狀結構,細胞膜結構變的復雜,黑色箭頭所指示的是T細胞活化后伸出的類似偽足的結構。從其剖面圖可以得出PHA活化后細胞高度為2.47μm,半高寬為5.47μm(圖3d)。從AFM高分辨的圖像可以看出,PHA活化的T細胞表面有豐富的顆粒狀團簇結構如圖3e和3f,團簇大小分布在100~500 nm。從 LFM模式圖(圖3g)可以看出,與靜息細胞(圖2g)相比,經PHA活化后細胞膜表面的摩擦力的分布變的不均勻,這與PHA活化后膜納米結構的變化相吻合,活化后細胞膜變的不均一。從AFM的定量數據可以得到PHA活化后T細胞膜表面顆粒大小在40~110 nm,呈正態分布,主要集中在80 nm左右,其中較大的顆粒粒徑達到155 nm如圖3h。從圖3i到3k可以看到SEA活化后細胞周圍同樣有豐富的蹼狀結構和偽足結構(圖中虛線和箭頭所示),從圖3l可以看出SEA活化后 T細胞的高度為 2.71μm,半高寬為5.31μm。從 AFM高分辨的圖像可以看出,SEA活化后膜納米結構變的復雜,表面起伏不平,與PHA活化的現象類似同樣有顆粒狀團簇的形成,團簇大小100 nm ~1μm(圖3m和3n),膜表面摩擦力的分布也變的不均勻(圖3o),組成團簇的顆粒大小在15~120 nm之間,呈正態分布,顆粒大小主要集中在75 nm左右,其中較大的顆粒達到134 nm(圖3p)。
為了對活化前后T細胞形態與膜納米結構進行定量對比分析,我們對關于細胞形態的各項物理參數做了統計分析,如圖4所示,這些物理參數主要包括:細胞體積、細胞直徑、細胞高度、膜表面平均粗糙度及膜表面納米顆粒高度分布等五個指標。統計結果直觀的表明:細胞經PHA和SEA刺激活化后,細胞的高度增加,細胞的直徑和體積也均增大,其中體積增加幅度較大(如圖4a和4b)。對于膜表面納米結構而言,其表面平均粗糙度略有增加但變化并不明顯,靜息的T細胞Ra為6 nm左右,PHA活化后Ra增加到8 nm左右,SEA活化后Ra增加到近10 nm,而膜表面顆粒的平均高度則明顯增大,靜息的T細胞表面顆粒高度為42 nm左右,PHA活化后顆粒高度增加到78 nm左右,SEA活化后顆粒高度增加到近91 nm左右(如圖4c所示),這與其超微結構的變化(圖2e到圖3e和圖3m)相吻合。需要說明的是經SEA活化后細胞體積、細胞直徑、細胞高度、膜表面平均粗糙度及膜表面納米顆粒高度這五項指標的增加幅度均大于PHA活化后的,這說明不同的刺激劑對T細胞的活化程度不同從而會導致不同的形態結構和膜納米結構。
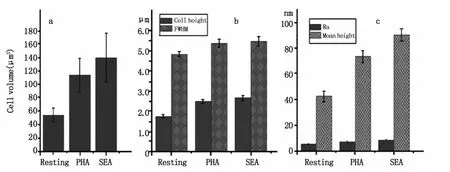
圖4 對活化前后T細胞的形貌及膜表面納米結構進行定量分析包括細胞體積,我們統計了其細胞體積(圖a),半高寬和細胞高度(圖b),以及膜納米結構變化的參數平均粗糙度(Ra)和顆粒平均高度(圖c)的變化Fig.4 Quantitative analysis of morphology and nanostructure about resting and activated T cells.A statistics of cell volume(a),cell height(b),full width at half maximum(FWHM)(b),average roughness(Ra)(c)and particle average height(c)was performed
2.3 不同外源刺激下T細胞膜納米力學特性的變化
T細胞的活化實驗采用PHA和SEA刺激T細胞活化。我們知道,植物凝集素(PHA)是一種從紅蕓豆中提取的有絲分裂原,作為一種低聚糖與蛋白質的復合物,植物凝集素有促進淋巴細胞有絲分裂、激活免疫細胞--淋巴細胞的作用。PHA刺激T細胞增殖分化產生大量效應T細胞和細胞毒T細胞;效應T細胞分泌產生大量細胞因子(如白介素-2、干擾素等)殺傷病毒[12]。超抗原(SEA)是一種蛋白激酶C的激活劑,可直接與 T細胞受體(T cell receptor,TCR)的某些Vβ區結合,不受主要組織相容性復合物(major histocompability complex,MHC)的限制,導致帶有特異性Vβ節段的 T細胞大量活化增[13]。
免疫是機體針對外源性物質的防御性反應。任何外源性物質首先侵入的是細胞膜,細胞膜已經被證明參與刺激性或者細胞毒性免疫反應,所以細胞膜的改變被認為是在分子水平上對細胞行為的影響[14]。我們前面的觀察結果表明了T細胞形貌及膜表面納米結構在經PHA和SEA刺激活化后發生改變,由于特異性膜分子的表達和分泌細胞因子等導致細胞表面的蛋白質等大分子明顯增多,從而引起細胞膜納米結構的相應變化。而細胞膜受到干擾或破壞,通過一定的途徑觸發細胞內的一系列生理生化反應,介導細胞粘著,即增強細胞間的粘連及細胞與基質的粘連,調節細胞的形狀和細胞骨架的結構。T細胞面對外源侵入,產生免疫應答,與靶細胞特異性結合,破壞靶細胞膜,直接殺傷靶細胞,其粘附力的大小直接決定著細胞免疫應答的能力。另一方面細胞膜表面蛋白等大分子含量的改變會導致細胞硬度的改變。細胞膜表面分子的變化可能會引起其力學特性的變化,這必然是與其免疫功能相適應的。以往的研究結果表明,細胞形態將會影響其機械性能[15,16],這表明,不同的形態則有著不同的機械性能。但是在AFM之前,人們還無法在納米尺度獲取細胞膜納米力學特性變化的參數[17],AFM作為獲得生物分子和細胞的納米結構和生物力學特性的強有力的工具,早已被證明是一種強有力的非破壞性的納米技術,AFM這一納米力感應器的出現,從而成為研究細胞動力學過程的新工具[18-21]。
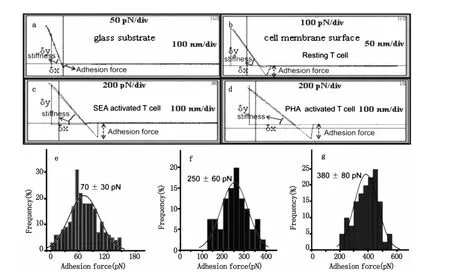
圖5 AFM針尖-細胞力測量結果。圖a是利用AFM探針獲得的玻璃表面非特異性作用的力曲線。圖b、圖c和圖d分別是AFM針尖和靜息的T細胞、SEA活化的細胞和PHA活化的細胞之間的非特異性的力曲線。圖e、圖f和圖g分別為靜息的、SEA活化的和PHA活化的T細胞膜表面粘附力的統計圖Fig.5 Measurement of the adhesion force of AFM tip-T cells,(a)Force-distance curve between AFM tip and glass surface,(b)Force-distance curve of resting T cells,(c,d)Force-distance curves of T cells which were activated by SEA and PHA for 48h,respectively.(e,f,g)Histograms of statistical results of surface adhesion force for resting groups,SEA groups and PHA groups,respectively
圖5a是針尖與玻璃表面的力曲線,玻璃表面與探針的作用力不明顯,從圖5b可以看出Resting T細胞表面與探針間具有明顯的粘附力。圖5c是SEA刺激48h后,針尖和T淋巴細胞表面的力曲線,與靜息的T細胞相比,SEA活化后粘附力增大;圖5d是PHA刺激后T淋巴細胞與針尖之間的力曲線,與靜息的T細胞和SEA活化的T細胞相比,PHA活化后細胞表面粘附力變得更大。通過定量分析可知AFM針尖與靜息的(圖5e)、SEA活化的(圖5f)和PHA活化的(圖5 g)T細胞之間的作用力分別為70 pN、250 pN和380 pN。T淋巴細胞在經過不同抗原刺激后,不但細胞形態和膜納米結構發生了變化,而且這種變化還伴隨著細胞膜納米機械性能的變化。不同的抗原刺激會導致粘附力的增加幅度不同,通過上面的分析可知PHA活化后T細胞表面的粘附力增加幅度較大。經抗原刺激活化后細胞表面的粘附力為靜息的細胞表面的粘附力的3-6倍。T細胞正是通過以上途徑,把與外源性物質作用后的變化表現在細胞微觀形貌上,這種變化進一步導致功能的變化,T細胞的這種變化形式可能使得其更加有效的與病原體相互作用從而進一步清除病原體。結合T淋巴細胞的超微形貌圖,靜息的T細胞膜表面較為平坦時,細胞的粘附力較小,當活化后隨著膜分子的表達,細胞膜成分團聚形成顆粒時粘附力則隨之增大。另一方面,前面的倒置顯微鏡圖1c到圖1f,顯示活化后細胞出現凝集現象,這種凝集作用依賴于LFA-1/ICAM-1的相互作用,靜息T淋巴細胞表面粘附分子LFA-1/ICAM-1結合較弱,T淋巴細胞在受到外來抗原刺激作用活化后,LFA-1/ICAM-1的結合顯著增強[27]。T細胞經PHA活化后粘附力較SEA活化后粘附力大,與前面的倒置顯微鏡所顯示的結果一致,即PHA活化后細胞凝聚團簇較SEA活化后細胞凝聚團簇大而且凝聚團簇多,這可能是PHA活化后LFA-1/ICAM-1的親和力更強的緣故。已有大量研究證明活化后細胞表面LFA-1/ICAM-1這兩種粘附分子的表達并沒有顯著增加,上述事實提示在T淋巴細胞活化后,粘附分子可能通過構型變化的方式,提高LFA-1/ICAM相互作用的親和力,從而提高活化淋巴細胞的粘附能力[22-24]。我們可以利用力曲線探針與樣品接觸部分的斜率變化來分析樣品表面的硬度變化,即δy/δx的比值越大,其硬度也越大。比較圖5a和5b可以看出玻璃表面的硬度較細胞表面的硬度大,與靜息的T細胞相比,發現SEA活化后(圖5c)和PHA活化后(圖5d)所測得的斜率要小于對靜息細胞所測得的斜率,因為曲線斜率與細胞硬度具有相關性[25],因此斜率變小意味著活化后T細胞的硬度變小,這可能與細胞在受到抗原刺激后的變形能力有關,如更容易伸出偽足等,同時細胞硬度減小有利于細胞的運動,從而更有利于與靶細胞等相互作用。
3 討論
細胞的超微結構與機械性能對細胞的生理學、病理學、藥理學過程起著重要的作用,如細胞表面顆粒及分子的分布,細胞的增殖、活化、凋亡、信號傳導等均與細胞的超微結構與機械性能相關。原子力顯微鏡這一科學研究工具不僅在細胞形態和膜納米結構研究中具有極大應用價值,而且在生物樣品,如細胞、生物大分子等納米力學測量等方面也具有無可比擬的優勢。
T細胞是免疫系統的重要組成部分,機體一旦受到病原體的攻擊,T細胞會迅速做出反應,從而導致其處于活化增殖狀態。我們首先利用PHA和SEA刺激活化T細胞,啟動機體多種免疫反應,并應用AFM的高空間分辨率比較了靜息、PHA活化和SEA活化的T淋巴細胞的超微形貌結構,并測量了細胞的納米機械性能。分析靜息的T細胞和活化的T細胞的形態結構和膜納米結構有利于進一步了解T細胞活化的機制和免疫突觸的形成以及整個免疫反應的過程。分析靜息的T細胞的AFM圖可以得知,靜息的T細胞膜表面較光滑(圖2a到2c),納米結構表明膜表面的分子排列緊密(圖2e和2f),測量表明膜表面平均粗糙度(Ra)分布為6.7 nm。靜息的細胞膜表面結構比較光滑、緊密,與之對應的LFM模式圖也表明靜息細胞的膜結構光滑的特點,其摩擦力的分布相對均勻(圖2g),這說明膜表面的各種生物大分子比較均勻的分布于膜表面,這與我們前面的成像結果相符合。雖然靜息的T細胞表面大部分地方摩擦力相同且較小,但是有的地方摩擦力要高于周圍的摩擦力,有的地方摩擦力低于周圍的摩擦力這是由于細胞表面顆粒高度的不同(圖2h)和細胞膜成分的不同引起的。與靜息的T細胞相比,經過PHA和SEA活化后,細胞的高度和體積明顯增大(圖4),這是T細胞經過刺激劑活化后增殖、分化而增大的表現。從其納米結構圖(圖3e和3m)可以看出活化后的T細胞表面布滿顆粒狀團簇(100 nm~1μm),這些微納結構域可能是在細胞活化與增殖的過程中,多種細胞因子和標志分子開始表達分泌,細胞表面的蛋白質等大分子明顯增多,T細胞表面分子會由疏散分布轉而聚集成簇從而形成免疫突觸[26],同時T細胞之間的相互作用和信息傳導也逐漸增加,細胞膜結構重排形成的具有其特殊功能的膜納米結構域,這對于免疫細胞的功能發揮是至關重要的。
經過PHA和SEA活化后的T細胞表面粘附力增大,是靜息的T細胞的3-6倍,結合其超微結構分析,活化后T細胞表面形成大小不一的顆粒狀物質,這些物質均為細胞表面蛋白和脂糖等的聚集物,細胞的這些形態變化引起其粘附力的變化;結合其功能分析,粘附力增大有利于加強T細胞與病原體的相互作用。另一方面活化后細胞硬度減小,這可能與細胞在受到抗原刺激后的變形能力有關,如更容易伸出偽足等,同時硬度減小有利于細胞的遷移,從而更有利于與靶細胞等相互作用從而進一步清除病原體。通過AFM的研究,可以進一步的了解T淋巴細胞形態變化與細胞行為之間的關系,為更好地理解T細胞的結構與功能提供了更多可視化的依據。
[1]MYERS M D,SOSINOWSKI T,DRAGONE L L,et al.Src-like adaptor protein regulates tcr expression on thymocytes by linking the ubiquitin ligase c-Cbl to the TCR complex[J].Nat Imm,2006,7(1):57-66.
[2]胡明鉛,蔡繼業.T細胞抗原識別與活化研究[J].生命科學研究,2008,12(3):190-192.HU Mingqian,CAI Jiye.Research on T cell antigen recognition and activation[J].Life Science Research,2008,12(3):190-192.
[3]SU Y T,MICHAEL L D.T-Cell Activation:A multidimensional signaling network[J].Curr Opin Cell Biol,2002,14(5):575-580.
[4]ZHONG L Y,LIAO W T,WANG X P,et al.Detection the specific marker of CD3 molecules of human peripheral blood t lymphocytes using SNOM and quantum Qots[J].Colloids Surf,A:Physicochem Eng Aspects,2008,313:642-646.
[5]廖問陶,鐘麗云,查慶兵,等.人外周血淋巴細胞體外刺激活化的原子力顯微鏡研究[J].激光雜志,2006,27(5):89-90.LIAO Wentao,ZHONG Liyun,CHA Qingbing,et al.Observation of the activation of human peripheral blood lymphocytes with atomic force microscopy[J].Laser Journal,2006,27(5):89-90.
[6]HUANG X,LI X,CHEN L,et al.Single-Molecule level binding force between collagen and collagen binding domain-growth factor conjugates[J].Biomaterials,2013,34:6139-6146.
[7]黃訓,金花,趙宏霞,等.原子力顯微鏡對中性粒細胞與K562細胞超微結構及機械性能的探測[J].分析測試學報,2009,28(11):1255-1259.HUANG Xun,JIN Hua,ZHAO Hongxia,et al.Ultrastructure and mechanical property of neutroph ilic granulocyte and leukem ia cellk562 detected by AFM[J].Journal of Instrumental Analysis,2009,28(11):1255-1259.
[8]GREGORY F,SARAH L,DAVID A,et al.Detection,localization,and conformational analysis of single polysaccharide molecules on live bacteria[J].ACS Nano,2008,2(9):1921-1928.
[9]MICHAEL L,DUSTIN A R.Reprograming t cells:the role of extracellular matrix in coordination of t cell activation and migration[J].Curr Opin Immunol,2001,13(3):286-290.
[10]EVANS E A,CALDERWOOD D A.Forces and bond dynamics in celladhesion[J]. Science, 2007, 316(5828):1148-1153.
[11]MALEK A M,IZUMO S.Control of endothelial cell gene expression by flow[J].J Biomechan,1995,28(12):1515.
[12]DONALD M,SIMONS E M,GARDNER P I.Sub-mitogenic phorbol myristate acetate co-stimulation rescues the pha induced activation of both na?ve and memory t cells cultured in the rotating-wall vessel bioreactor[J]. Cell Biology International,2009,33(8):882-886.
[13]CRAIG T,MORITA H M,LI J G,et al.Superantigen Recognition by γδ T Cells:SEA recognition site for human Vγ2 T cell receptors[J].Immunity,2001,14(3):331-344.
[14]ROOZEMOND R C,BONAVIDA B.Effect of altered membrane fluidity on NK cell-mediated cyto-toxicity selective-inhibition of the recognition or post recognition events in the cytolytic pathway of NK cells[J].J Immunol,1985,134(4):2209-2214.
[15]ADDAE-MENSAH K A,WIKSWO J P.Measurement techniques for cellular biomechanics in vitro[J].Exp Biol Med,2008,233(7):792-809.
[16]SCHMITZ J,GOTTSCHALK K E.Mechanical regulation of cell adhesion[J].Soft Matter,2008,4(7):1373-1387.
[17]ZHANG X,WOJCIKIEWICZ E,MOY V T.Force spectroscopy of the leukocyte function-associated antigen-1/intercellular adhesion molecule-1 interaction[J].Biophys J,2002,83:2270-2279.
[18]DUFRENE Y F.Atomic force microscopy,a powerful tool in microbiology[J].J Bact,2002,184(19):5205-5213.
[19]DUFRENE Y F.Using nanotechniques to explore microbial surfaces[J].Nat Rev Microbiol,2004,2(6):451-460.
[20]GREENLEAF W J,WOODSIDE M T,BLOCK S M.High-resolution,single-molecule measurements of biomolecular motion[J].Annu Rev Biophys Biomol Stru,2007,36:171-190.
[21]SURESH S.Biomechanics and biophysics of cancer cells[J].Acta Biomater,2007,3(4):413-438.
[22]LUPHER M L,HARRIS E A S,BEALS C R,et al.Cellular activation of leukocyte function-associated antigen-1 and its affinity are regulated at the i domain allosteric site[J].J Immunol,2001,167(3):1431-1439.
[23]SCOTT A T,SEETHARAMA D S,SIAHAAN T J,et al.Linear and cyclic lfa-1 and icam-1peptides inhibit t cell adhesion and function[J].Peptides,2000,21(8):1161-1167.
[24]ZHANG W Y,SHAO Y,FANG D Y,et al.Negative regulation of t cell antigen receptor-mediated crk-l-c3g signaling and cell adhesion by Cbl-b[J].J Biol Chem,2003,278(26):23978-23983.
[25]ABU-LAIL N I,CAMESANO T A.Role of ionic strength on the relationship of biopolymer conformation,DLVO contributions,and steric interactions to bioadhesion of pseudomonas putida KT2442[J].Biomacromolecules,2003,4:1000-1012.
[26]MICHELLE K,JOHANNES B,HUPPA M A,et al.Linking molecular and cellular events in t-cell activation and synapse formation[J].Sem Immunol,2003,15(6):307-315.
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
