產(chǎn)耐高溫木聚糖酶菌株的篩選及其產(chǎn)酶條件優(yōu)化
2013-04-23 02:07:42孫明哲鄭宏臣孫君社裴海生劉逸寒路福平
中國釀造 2013年8期
孫明哲,鄭宏臣,孫君社,3,裴海生,劉逸寒,張 璟,韓 楊,路福平*
(1.天津科技大學(xué) 生物工程學(xué)院 工業(yè)發(fā)酵微生物教育部重點(diǎn)實(shí)驗(yàn)室 工業(yè)酶國家工程實(shí)驗(yàn)室 天津市工業(yè)微生物重點(diǎn)實(shí)驗(yàn)室,天津 300457;2.農(nóng)業(yè)部規(guī)劃設(shè)計研究院,北京 100125;3.中國農(nóng)業(yè)大學(xué)食品科學(xué)與營養(yǎng)工程學(xué)院,北京 100083)
木聚糖是被子植物(闊葉木和和本科)細(xì)胞壁中半纖維素的主要成分,其含量僅次于纖維素,是一種豐富的可再生資源[1]。作為一種異質(zhì)性雜多糖,其主鏈?zhǔn)怯蒁-木糖以β-1,4-糖苷鍵連接而成,其側(cè)鏈由種類各異的糖基側(cè)鏈組成[2]。內(nèi)切-1,4-β-D-木聚糖酶作用于木聚糖主鏈,是木聚糖降解酶系中的關(guān)鍵酶,通常被簡稱為木聚糖酶[3]。木聚糖酶的來源相當(dāng)廣泛,已報道產(chǎn)木聚糖酶的微生物主要有細(xì)菌、放線菌、真菌、木霉、青霉、曲霉等,當(dāng)前絕大多數(shù)商用木聚糖酶來源于霉菌,主要是由于霉菌微生物產(chǎn)酶量相對較高,但霉菌發(fā)酵周期長,并且大部分霉菌木聚糖酶最適作用條件偏于中性,使其在造紙、飼料、食品等工業(yè)的極端環(huán)境下應(yīng)用受到一定限制[4-6]。而近年來報道的細(xì)菌木聚糖酶在耐極端條件方面有較強(qiáng)的優(yōu)勢,具有較好的開發(fā)前景[7]。
目前已報道的產(chǎn)木聚糖酶的細(xì)菌以芽孢桿菌居多。2012年,CHARBONNEAU DM等[8]在加拿大的肥料中篩選出多株產(chǎn)耐熱木聚糖酶的細(xì)菌,其最適作用溫度為60℃~65℃;2013年,ANAND A等[9]從印度的高溫堆肥中篩選出一株嗜熱脫氮芽孢桿菌(Geobacillus thermodenitrificansTSAA1),其所產(chǎn)木聚糖酶的最適溫度為70℃。我國在產(chǎn)木聚糖酶細(xì)菌的研究方面也有顯著成果,鄭宏臣等[10]從造紙廠周邊土壤中分離純化到一株產(chǎn)堿性木聚糖酶的短小芽孢桿菌(Bacillus pumilusG1-3),并對其酶學(xué)性質(zhì)進(jìn)行分析,其最適反應(yīng)pH值和溫度分別為8.0和55℃;梁方方等[11]從長期堆放青貯飼料的土壤中分離到一株枯草芽孢桿菌X7,對其酶學(xué)性質(zhì)的初步研究顯示該酶最適pH值和溫度分別為6.0和50℃;陳學(xué)敏等[12]從云南騰沖熱泉水樣中分離得到一株產(chǎn)木聚糖酶的菌株,并鑒定為地芽孢桿菌(Geobacillussp.PZH1),其木聚糖酶的最適pH值和最適溫度分別為7.0和70℃;孫振濤等[13]從土壤中分離到一株產(chǎn)木聚糖酶的類芽孢桿菌(Paenibacillus campinasensis),其產(chǎn)木聚糖酶的最適pH值和最適溫度分別為7.0和60℃。
本研究以篩選耐熱木聚糖酶的產(chǎn)生菌為目的,從北京市順義區(qū)的養(yǎng)牛場堆肥中獲得一株可產(chǎn)耐熱木聚糖酶的細(xì)菌,初步鑒定為類芽孢桿菌。對其木聚糖酶的酶學(xué)性質(zhì)進(jìn)行了初步研究,并優(yōu)化了其發(fā)酵產(chǎn)酶的培養(yǎng)條件。同時本研究也進(jìn)一步豐富了產(chǎn)木聚糖酶菌的菌種資源。
1 材料與方法
1.1 材料與試劑
牛糞樣品:采集自北京市順義區(qū)趙全營鎮(zhèn)豹房村西北奶牛養(yǎng)殖場。
選擇培養(yǎng)基:稻殼木聚糖1.00%,NH4NO30.50%,MgSO40.03%,NaCl 0.50%,K2HPO40.20%,(NH4)2SO40.10%,酵母粉0.03%,瓊脂2.00%,pH值自然;種子培養(yǎng)基:牛肉粉0.50%,蛋白胨1.00%,NaCl 0.50%,pH值自然;基礎(chǔ)產(chǎn)酶培養(yǎng)基:麩皮4.00%,蛋白胨1.00%,K2HPO40.50%,MgSO40.05%,NaCl 0.5%,pH值自然。
NaCl、NaOH、MgSO4、NH4NO3、K2HPO4、Na2HPO4、(NH4)2SO4、3-5-二硝基水楊酸(DNS)、苯酚、四水合酒石酸鉀鈉、偏重亞硫酸鈉、尿素、檸檬酸均為國產(chǎn)分析純;稻殼木聚糖購自天津市光復(fù)精細(xì)化工研究所;樺木木聚糖購自Sigma 公司;麩皮、玉米芯、玉米桿經(jīng)粉碎后過40目篩。
1.2 儀器與設(shè)備
YT-CJ-2ND型潔凈工作臺、TU-1901雙光束紫外可見分光光度計:北京普析通用儀器有限責(zé)任公司;HZQ-T160型恒溫培養(yǎng)箱:哈爾濱東聯(lián)電子技術(shù)開發(fā)有限公司法;BS 200 S型分析天平:北京賽多利斯天平有限公司。
1.3 方法
1.3.1 初篩方法
稱1g牛糞樣品于100mL無菌水中,振蕩搖勻制得菌懸液。取1mL菌懸液加入到20mL滅菌的富集培養(yǎng)基中,于80℃水浴加熱處理30min,以殺死不耐熱的微生物營養(yǎng)細(xì)胞;然后置于50℃搖床上,培養(yǎng)2d。富集培養(yǎng)液經(jīng)適當(dāng)稀釋后涂布于篩選平板培養(yǎng)基,置50℃恒溫培養(yǎng)箱培養(yǎng)1d~2d,選取能產(chǎn)生透明圈的單菌落進(jìn)一步劃線純化并編號,即木聚糖酶產(chǎn)生菌,作為進(jìn)一步的復(fù)篩菌種。
1.3.2 復(fù)篩
將菌種活化16h后接入基礎(chǔ)產(chǎn)酶培養(yǎng)基中進(jìn)行搖瓶復(fù)篩,搖瓶使用100mL三角瓶裝液50mL,180r/min、50℃,取72h發(fā)酵液4000r/min離心10min制得粗酶液,測其酶活力,選取酶活較高者保菌。
1.3.3 菌種鑒定
細(xì)胞形態(tài)觀察參見微生物學(xué)試驗(yàn)技術(shù)[14]。16S rDNA序列分析:以產(chǎn)酶菌為模板,應(yīng)用16S rDNA通用引物27F(AGAGTTTGATCMTGGCTCAG)和1492R(TACGGYTA CCTTGTTACGACTT),通過菌落PCR擴(kuò)增產(chǎn)酶菌株的16S rDNA序列,PCR擴(kuò)增程序:95℃、5min;94℃、45s,55℃、45s,72℃、90s,30個循環(huán);72℃延伸10min。PCR結(jié)果送北京三博遠(yuǎn)志生物技術(shù)有限公司測序,測序結(jié)果在GenBank中進(jìn)行比對。
1.3.4 Plackett-Burman試驗(yàn)
首先選擇麩皮、玉米芯、玉米桿、稻殼木聚糖4種碳源及蛋白胨、牛肉膏、酵母浸粉、尿素、(NH4)2SO4、NH4NO36種氮源進(jìn)行單因素試驗(yàn),以確定最適的發(fā)酵產(chǎn)酶碳、氮源。
選取碳源濃度、氮源濃度、K2HPO4濃度、MgSO4濃度、NaCl濃度、溫度、搖床轉(zhuǎn)速、裝液量、接種量、發(fā)酵初始pH值這10個因素進(jìn)行Plackett-Burman試驗(yàn)[15](因素水平設(shè)計見表1)以確定影響發(fā)酵產(chǎn)酶重要因素。
1.3.5 響應(yīng)面法優(yōu)化發(fā)酵條件
根據(jù)Plackett-Burman試驗(yàn)的結(jié)果,確定影響發(fā)酵產(chǎn)酶的3個主要因素A、B、C,依據(jù)Box-Behnken中心組合設(shè)計原理[16],設(shè)計3因素3水平試驗(yàn),共選取17個試驗(yàn)點(diǎn),包含12個析因點(diǎn),5個零點(diǎn)重復(fù),用以估計試驗(yàn)誤差。試驗(yàn)因素水平設(shè)計見表2。
1.3.6 木聚糖酶活力測定
木聚糖酶活力測定使用DNS法[7,17]。木聚糖酶活力單位的定義為:以1%的可溶性樺木木聚糖為底物,在酶的最適反應(yīng)條件下,每分鐘分解木聚糖生成1mol木糖所需酶量為一個酶活力單位(IU)。
2 結(jié)果與討論
2.1 木聚糖酶產(chǎn)生菌的篩選
試驗(yàn)通過高溫培養(yǎng)透明圈法從牛糞樣品中篩選出木聚糖酶產(chǎn)生菌,其中選取28株產(chǎn)生透明圈較大的進(jìn)行搖瓶發(fā)酵復(fù)篩,選取在高溫條件下酶活相對最高的菌株NF1,并對其進(jìn)行反復(fù)發(fā)酵驗(yàn)證后,發(fā)現(xiàn)菌株NF1在高溫條件下產(chǎn)木聚糖酶較高且穩(wěn)定性較好,該菌株發(fā)酵后制得的初酶液對樺木木聚糖底物的最適催化溫度及pH值分別為60℃和6.0(見圖1、圖2),并且在高溫條件下酶活可長時間保持穩(wěn)定,說明菌株NF1可產(chǎn)耐高溫木聚糖酶。
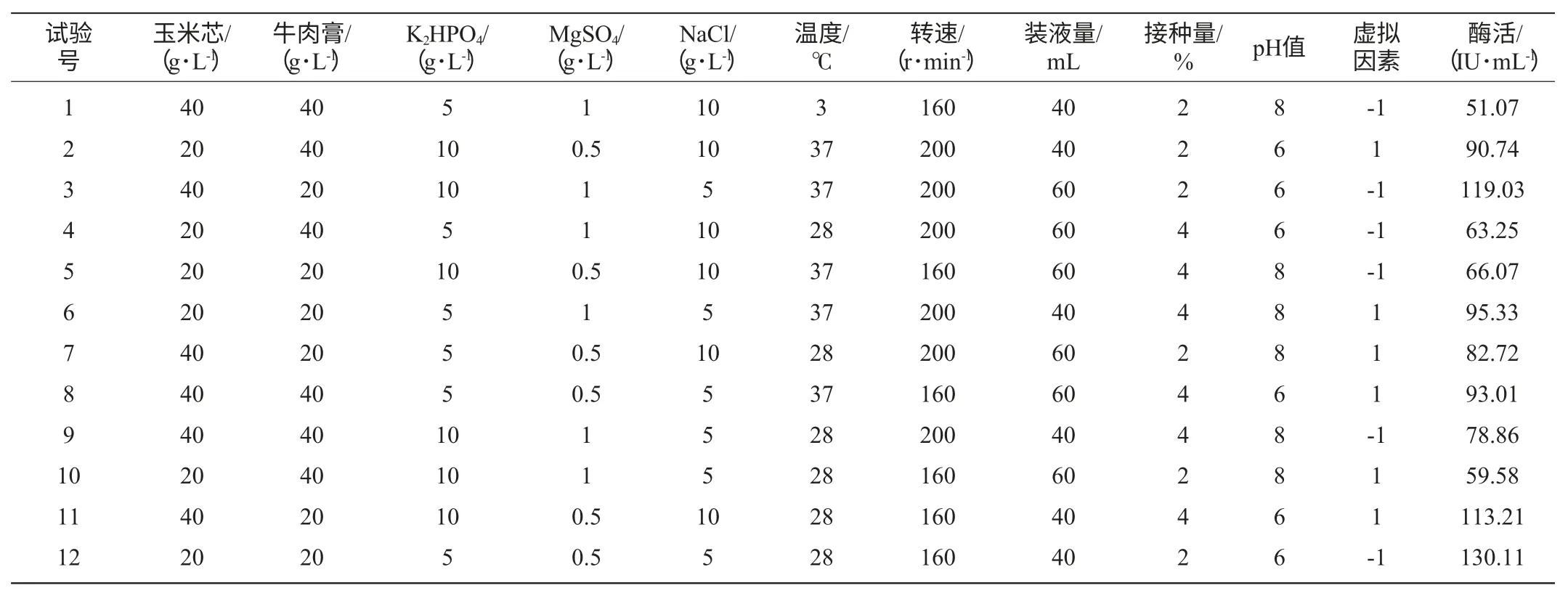
表1 Plackett-Burman試驗(yàn)設(shè)計及結(jié)果Table 1 Design and result of Plackett-Burman experiment
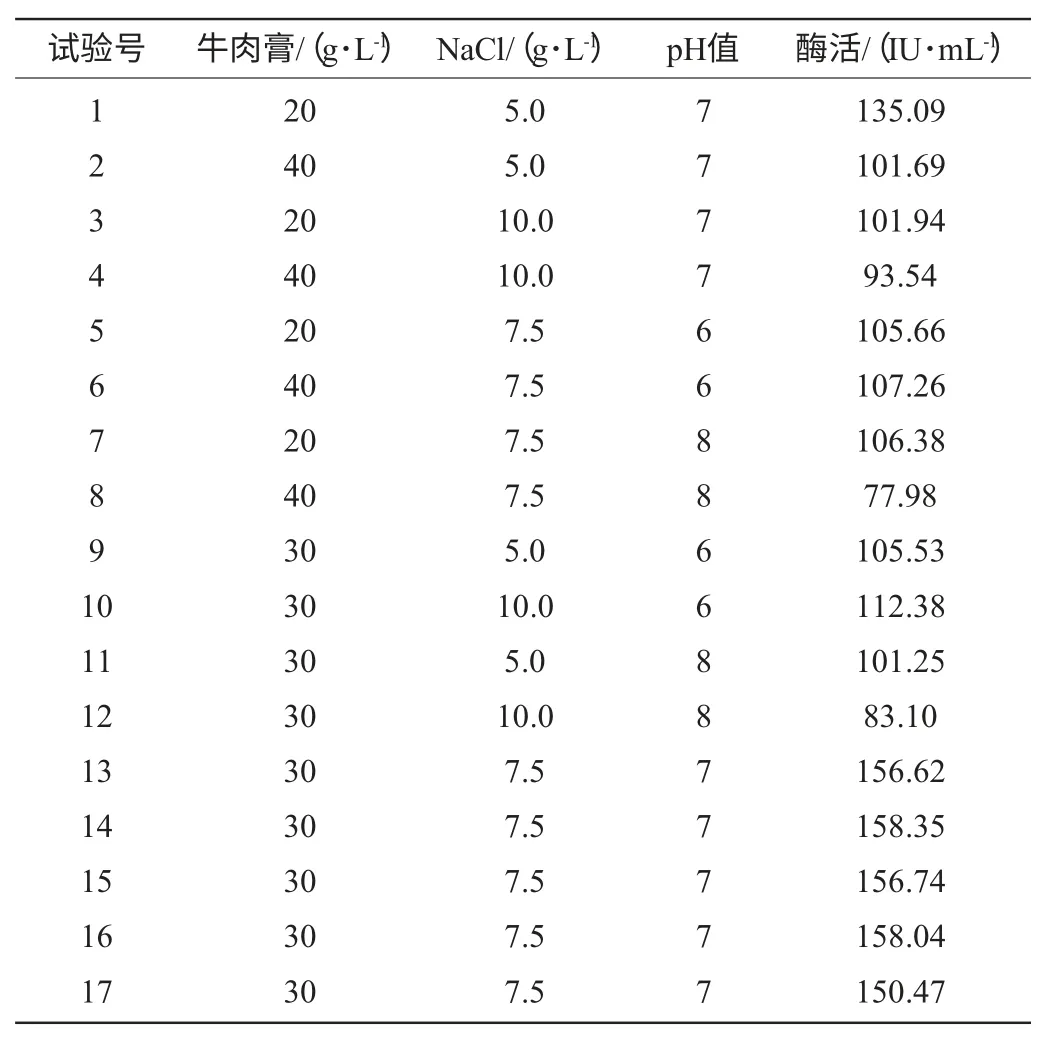
表2 Paenibacillus sp.NF1發(fā)酵產(chǎn)酶的Box-Behnken試驗(yàn)設(shè)計及結(jié)果Table 2 Experiment design and results of the Box-Behnken design for xylanase production
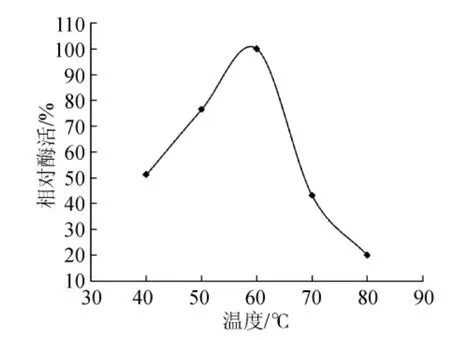
圖1 最適催化溫度Fig.1 The optimal catalytic temperature
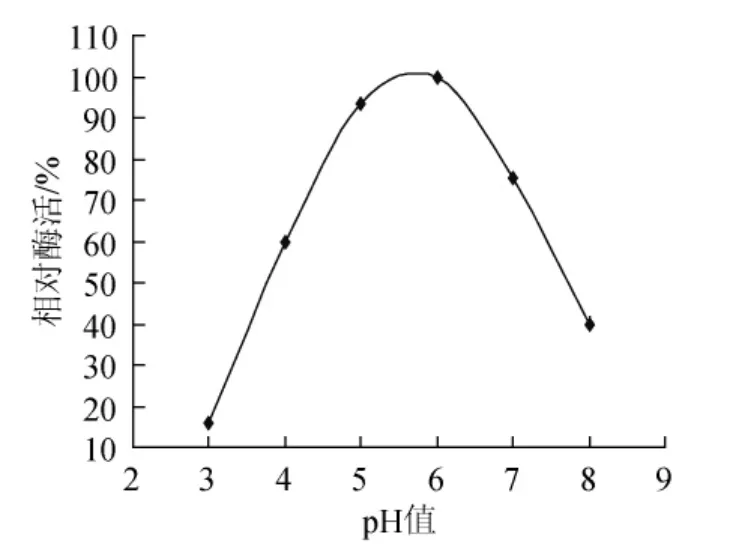
圖2 最適催化pH值Fig.2 The optimal catalytic pH value
2.2 菌種鑒定
菌株NF1的菌體形態(tài)呈桿狀,革蘭氏染色陽性(圖3);16S rDNA序列比對結(jié)果顯示該菌屬于類芽孢桿菌屬,命名為類芽孢桿菌(Paenibacillussp.)NF1。

圖3 菌株NF1的革蘭氏染色圖Fig.3 Gram’s staining of NF1
2.3 菌株NF1發(fā)酵產(chǎn)酶的Plackett-Burman試驗(yàn)
試驗(yàn)首先對Paenibacillussp.NF1發(fā)酵產(chǎn)酶的最佳碳、氮源進(jìn)行了考察,結(jié)果見圖4、圖5。當(dāng)使用稻殼木聚糖為碳源誘導(dǎo)產(chǎn)酶時發(fā)酵液酶活最高,但使用玉米芯為碳源時發(fā)酵液酶活與使用稻殼木聚糖相差不大,并明顯高于使用麩皮和玉米桿的發(fā)酵液酶活,因此考慮到經(jīng)濟(jì)因素這里選擇玉米芯為最佳碳源。氮源對發(fā)酵產(chǎn)酶的影響不是很大,選取酶活最高者牛肉膏為最佳氮源。確定最佳碳、氮源后,依據(jù)Plackett-Burman試驗(yàn)原理設(shè)計了試驗(yàn),用以判定影響Paenibacillussp.NF1發(fā)酵產(chǎn)酶的主要因素,試驗(yàn)設(shè)計見表1。使用Design-Expert軟件進(jìn)行試驗(yàn)結(jié)果的分析,結(jié)果見表3。
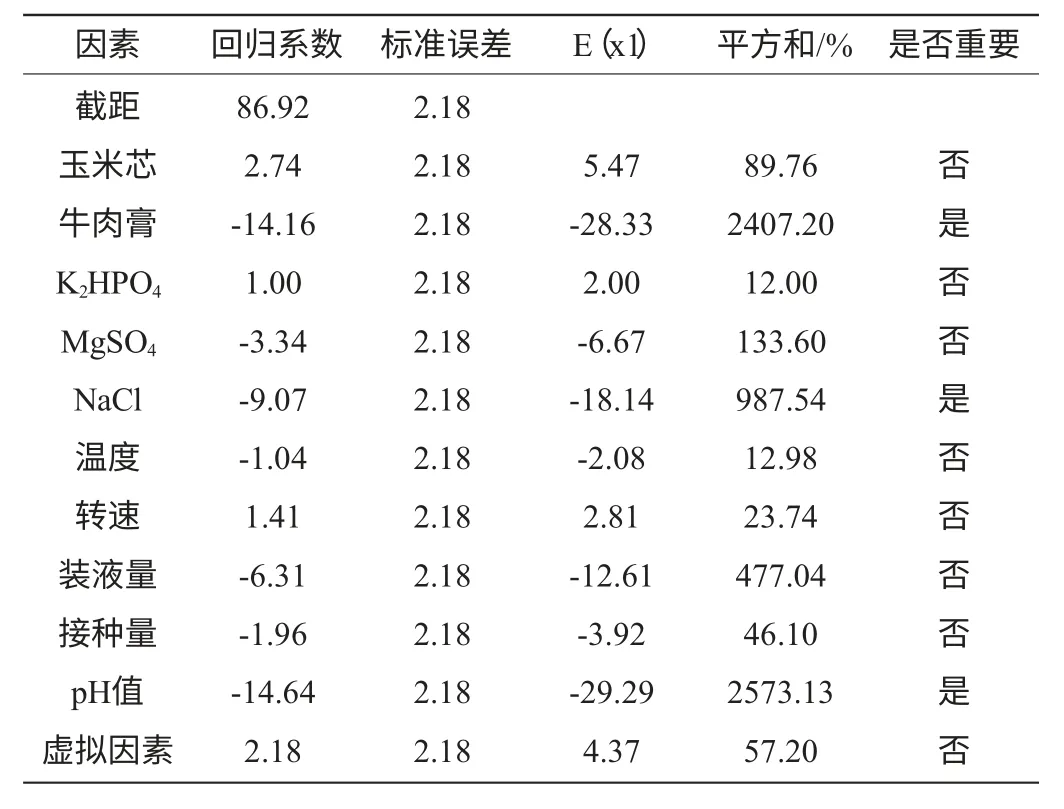
表3 偏回歸系數(shù)及因素的顯著性分析Table 3 Partial regression coefficients and analyses of their significance
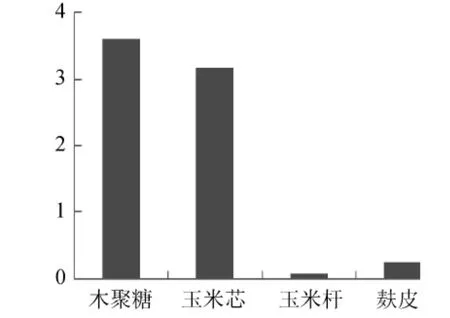
圖4 不同碳源對Paenibacillus sp.NF1發(fā)酵產(chǎn)酶的影響Fig.4 Effect of different carbon source on Paenibacillus sp.NF1 xylnanse enzyme production
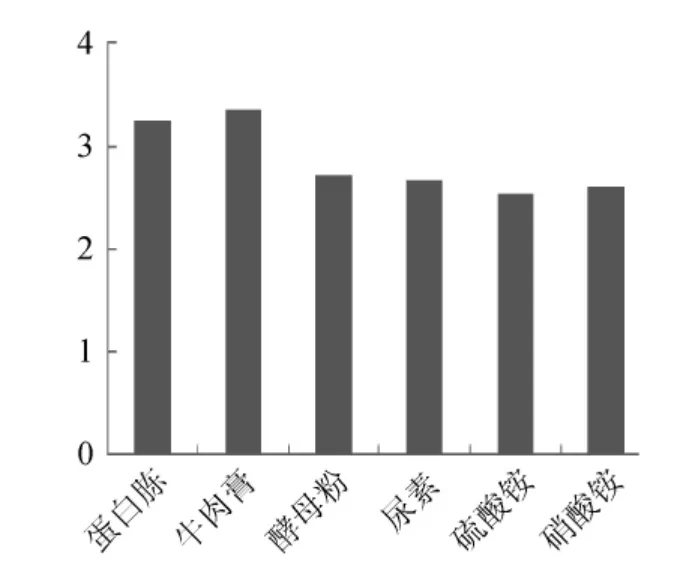
圖5 不同氮源對Paenibacillus sp.NF1發(fā)酵產(chǎn)酶的影響Fig.5 Effect of different nitrogen source on Paenibacillus sp.NF1 xylnanse enzyme production and response values
以牛肉膏濃度因素為例,其偏回歸系數(shù)為-14.16、標(biāo)準(zhǔn)誤差為2.18、影響水平E(牛肉膏g/L)=-28.33,表明牛肉膏濃度對發(fā)酵產(chǎn)酶的影響為負(fù)效應(yīng),因此,在后續(xù)試驗(yàn)中應(yīng)降低其水平;牛肉膏濃度的平方和百分比(contribution of SS)SS%=2407.20,明顯高于玉米芯濃度、K2HPO4濃度、MgSO4濃度、溫度、搖床轉(zhuǎn)速、裝液量、接種量等因素,因此顯著性分析結(jié)果為重要。由表3可明顯看出,牛肉膏濃度、NaCl濃度和發(fā)酵初始pH值為3個影響Paenibacillussp.NF1發(fā)酵產(chǎn)酶的主要因素,其影響值分別為2407.20%、987.54%、2573.13%,而虛擬因素對發(fā)酵產(chǎn)酶的影響值較低(57.20%)表明了該線性模型的適用性。
Plackett-Burman試驗(yàn)方差分析模型的Prob(P)>F值,為0.0006,表明所得回歸方程達(dá)極顯著水平,即該模型在被研究的整個回歸區(qū)域擬合的很好;復(fù)相關(guān)系數(shù)(R2)=0.8750,說明相關(guān)性較好;校正相關(guān)系數(shù)(Adj R2)=0.8281,表明82.81%的試驗(yàn)數(shù)據(jù)的變異性可用此回歸模型來解釋;通常情況下變異系數(shù)(CV)越低,試驗(yàn)的可信度和精確度越高,CV=11.88%,表示PB試驗(yàn)的可信度和精確度較好;精密度(Adeq Precision)是有效信號與噪聲的比值,大于4.0視為合理,本試驗(yàn)精密度為12.712(表4)。
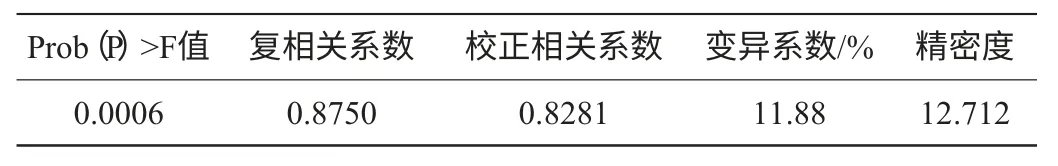
表4 Plackett-Burman試驗(yàn)方差分析Table 4 Analyses of variance for Plackett-Burman experiment
2.4 Paenibacillus sp.NF1發(fā)酵產(chǎn)酶條件的響應(yīng)面分析
根據(jù)Plackett-Burman試驗(yàn)結(jié)果得出牛肉膏濃度、NaCl濃度、發(fā)酵初始pH值為影響Paenibacillussp.NF1發(fā)酵產(chǎn)酶的3個主要因素,按照Box-Behnken設(shè)計每個因素取3個水平進(jìn)行試驗(yàn),3因素3水平共17組試驗(yàn),方案與結(jié)果見表2。
根據(jù)表2的試驗(yàn)結(jié)果,使用Design-Expert軟件進(jìn)行方差分析和二次回歸擬合試驗(yàn)數(shù)據(jù),回歸方程變量分析見表5。
以木聚糖酶活力(R1)為響應(yīng)面值,以A(牛肉膏濃度)、B(NaCl濃度)、C(發(fā)酵初始pH值)為自變量,擬合得到多元二次回歸方程:R1=156.4-8.75A-6.57B-7.77C+6.25AB-7.50AC-6.25BC-24.61A2-23.36B2-32.11C2
由回歸方程系數(shù)顯著性檢驗(yàn)及方差分析可知,模型試驗(yàn)擬合良好,模型的p<0.0001<0.05,表明該試驗(yàn)?zāi)P惋@著;失擬項(xiàng)p=0.0858>0.05,說明方程對試驗(yàn)的擬合度較好,此方法可靠。從表5中看到各因素對Paenibacillussp.NF1發(fā)酵產(chǎn)酶的影響不同,A、B、C的影響均為極顯著;AB、AC、BC對發(fā)酵產(chǎn)酶的效應(yīng)顯著;A2、B2、C2也均對發(fā)酵產(chǎn)酶存在極顯著影響。表明試驗(yàn)因子對響應(yīng)值存在非線性關(guān)系,其中二次項(xiàng)與響應(yīng)值也存在較大關(guān)系。模型的R2=0.9888,說明回歸方程的擬合程度良好,失擬性較小。
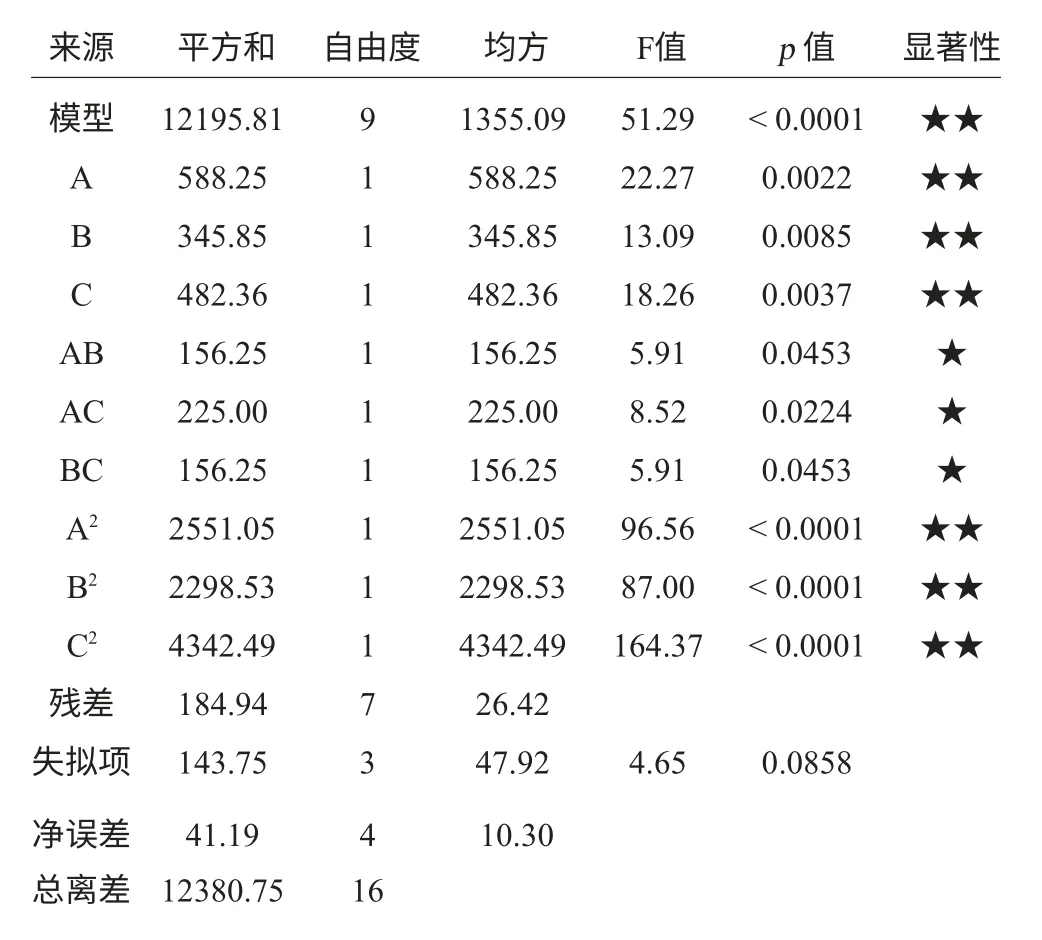
表5 回歸方程變量分析Table 5 Variance analysis of regression equation
由圖6可知,在發(fā)酵培養(yǎng)基初始pH值(pH=7.0)一定的條件下,木聚糖酶活力隨NaCl濃度和牛肉膏濃度的升高而先升高后降低,兩者交互作用顯著,單因素效應(yīng)相當(dāng)。
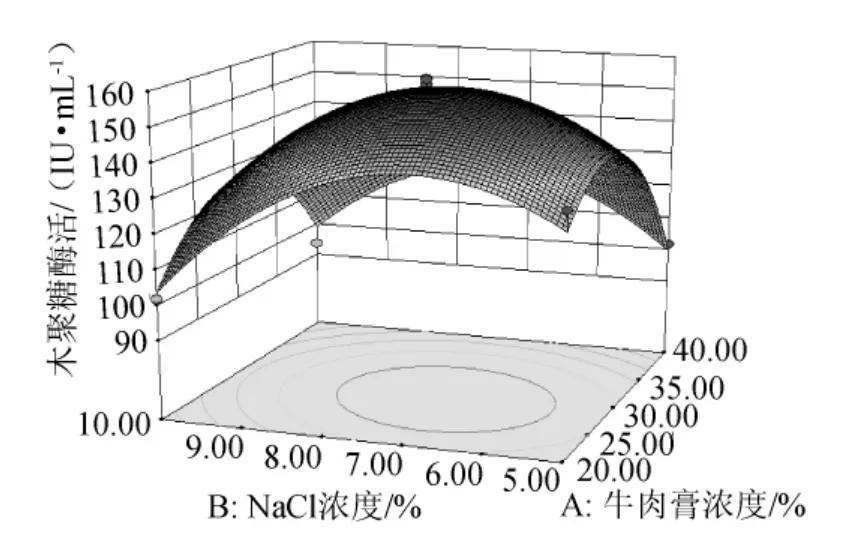
圖6 牛肉膏濃度和NaCl濃度交互作用的響應(yīng)面圖Fig.6 Response surface map of the combined effects between concentration of beef extract and NaCl
由圖7可知,在NaCl濃度一定的條件下,木聚糖酶活力隨牛肉膏濃度和培養(yǎng)基初始pH值的升高而呈現(xiàn)先升高后降低的趨勢,兩者交互作用顯著,且培養(yǎng)基初始pH值對木聚糖酶活力的影響比牛肉膏顯著。
由圖8可知,在牛肉膏濃度一定的條件下,木聚糖酶活力隨NaCl濃度和培養(yǎng)基初始pH值的增加而呈想升高后降低的趨勢,二者交互作用顯著,單因素效應(yīng)相當(dāng)。
求解回歸方程可得Paenibacillussp.NF1發(fā)酵產(chǎn)木聚糖酶的最優(yōu)條件為牛肉膏28.19g/L,NaCl 7.12g/L,pH 值為6.92,此時模型預(yù)測的產(chǎn)酶最大值為157.653U/mL。對模型進(jìn)行驗(yàn)證(取牛肉膏28.20g/L,NaCl 7.10g/L,pH值為6.90),設(shè)計3組平行,測得木聚糖酶酶活為160.6IU/mL,與預(yù)測酶活值偏差1.83%,無顯著差異。經(jīng)優(yōu)化后Paenibacillussp.NF1的產(chǎn)酶量為原始培養(yǎng)條件的51.4倍。
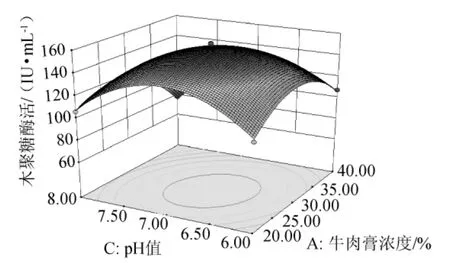
圖7 牛肉膏濃度和培養(yǎng)基初始pH值交互作用的響應(yīng)面圖Fig.7 Response surface map of the combined effects between concentration of beef extract and initial pH value
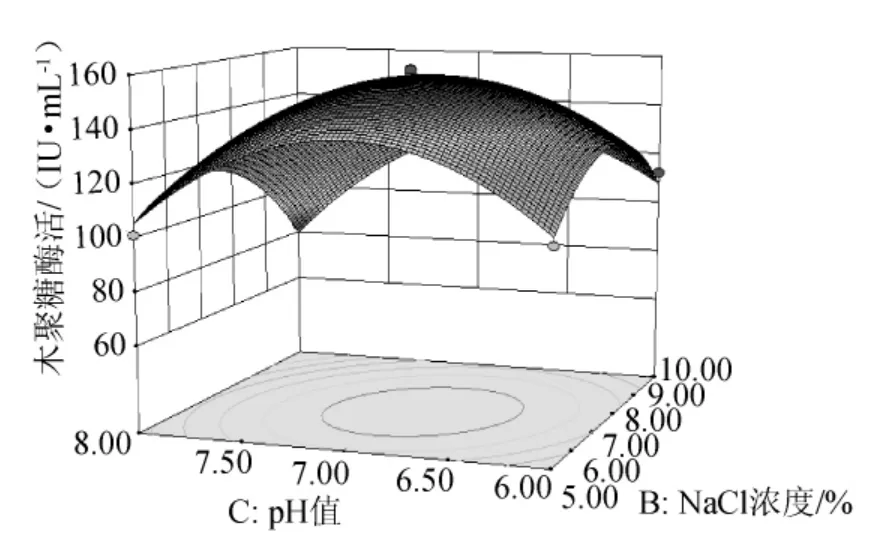
圖8 NaCl濃度和培養(yǎng)基初始pH值交互作用的響應(yīng)面圖Fig.8 Response surface map of the combined effects between concentration of NaCl and initial pH value
3 結(jié)論
利用透明圈法從牛糞中篩選得到一株高產(chǎn)耐熱木聚糖酶的菌株P(guān)aenibacillussp.NF1,其最適催化溫度和pH值分別是60℃和6.0。應(yīng)用響應(yīng)面法對該菌發(fā)酵產(chǎn)木聚糖酶條件進(jìn)行了優(yōu)化,確定其最適發(fā)酵培養(yǎng)基組成為玉米芯20g/L、牛肉膏28.2g/L、K2HPO45g/L、MgSO40.5g/L、NaCl7.1g/L、培養(yǎng)基初始pH值為6.9;最適發(fā)酵條件為裝液量100mL錐形瓶裝液40mL、搖床轉(zhuǎn)速160r/min、接種量2%、溫度28℃。此條件下發(fā)酵72h,木聚糖酶活力可達(dá)160.6IU/mL,與預(yù)測結(jié)果一致,為原始發(fā)酵條件產(chǎn)酶量的51.4倍。
[1]GREGORY ACE,O’CONNELL AP,BOLWELL PG.Xylans[J].Biotechnol Genet Eng,1998,15(4):439-455.
[2]丁長河.鏈霉菌高產(chǎn)木聚糖酶及其酶學(xué)性質(zhì)的研究[D].北京:中國農(nóng)業(yè)大學(xué)博士論文,2003.
[3]BEG QK,KAPOOR M,MAHAJAN L,et al.Microbial xylanases and their industrial applications:a review[J].Appl Microbiol Biot,2001,56(3-4):326-338.
[4]SUNNA A,ANTRANIHIAN G.Xylanolytic enzymes from fungi and bacteria[J].Crit Rev Biotechnol,1997,17(1):39-67.
[5]SHI P,QIU Z,BAI Y,et al.A new xylanase fromStreptomyces megasporusDSM 41476 with high yield of xylobiose[J].World J Microb Biot,2012,28(2):687-692.
[6]MENON G,MODY K,KESHRI J,et al.Isolation,purification,and characterization of haloalkaline xylanase from a marineBacillus pumilusstrain,GESF-1[J].Biotechnol Bioproc Eng,2011,15(6):998-1005.
[7]ZHENG HC,LIU XG,HAN Y.Isolation,purification and characterization of a thermostable xylanase from a novel strainPaenibacillus campinasensisG1-1[J].J Microbiol Biotechn,2012,22(7):930-958.
[8]CHARBONNEAU DM,MEDDEB-MOUELHI F,BOISSINOT M,et al.Identification of thermophilic bacterial strains producing thermotoleranthydrolytic enzymes from manure compost[J].Indian J Microbiol,2012,52(1):41-47.
[9]ANAND A,KUMAR V,SATYANARAYANA T.Characteristics of thermostable endoxylanase and β-xylosidase of the extremely thermophilic bacteriumGeobacillus thermodenitrificansTSAA1 and its applicability in generating xylooligosaccharides and xylose from agroresidues[J].Extremophiles,2013,17(3):357-366.
[10]鄭宏臣,劉逸寒,劉曉光,等.堿性木聚糖酶產(chǎn)生菌的篩選、XynG1-3 基因克隆表達(dá)及酶學(xué)性質(zhì)研究[J].生物技術(shù)通報,2012(10):106-113.
[11]梁方方,莫 毅,楊 琳.木聚糖酶產(chǎn)生菌的篩選鑒定及酶學(xué)性質(zhì)研究[J].飼料研究,2011(3):45-47.
[12]陳學(xué)敏,劉培培,張 波.一株產(chǎn)木聚糖酶嗜熱菌的鑒定及酶學(xué)性質(zhì)[J].微生物學(xué)通報,2011,38(2):151-156.
[13]孫振濤,劉建軍,趙祥穎.一株產(chǎn)木聚糖酶菌株的分離、鑒定及其酶學(xué)特性研究[J].生物技術(shù),2007,17(4):74-77.
[14]杜連祥,路福平.微生物學(xué)試驗(yàn)技術(shù)[M].北京:中國輕工業(yè)出版社,2008.
[15]孟 濤.關(guān)于Plackett-Burman 飽和設(shè)計數(shù)據(jù)分析方法的比較研究[D].上海:華東師范大學(xué)碩士論文,2008.
[16]黃新仁.響應(yīng)面法在生物過程優(yōu)化中的應(yīng)用[D].長沙:湖南大學(xué)碩士論文,2011.
[17]BAILEY MJ,BIELY P,POUTANEN K.Interlaboratory testing of methods for assay of xylanase activity [J].J Biotechnol,1992,23(3):257-270.
