嗜酸氧化亞鐵硫桿菌中鋅離子轉運基因的鑒定與分析
2012-11-03 03:32:10侯冬梅王洋洋張云靜劉代剛吳學玲
中國有色金屬學報 2012年5期
侯冬梅,苗 博,王洋洋,張云靜,劉代剛,吳學玲
(1. 中南大學 資源加工與生物工程學院,長沙 410083;2. 中南大學 教育部生物冶金重點實驗室,長沙 410083)
嗜酸氧化亞鐵硫桿菌中鋅離子轉運基因的鑒定與分析
侯冬梅1,苗 博1,王洋洋1,張云靜1,劉代剛1,吳學玲2
(1. 中南大學 資源加工與生物工程學院,長沙 410083;2. 中南大學 教育部生物冶金重點實驗室,長沙 410083)
鑒定了Acidithiobacillus ferrooxidans DC (At. ferrooxidans DC)中與鋅離子轉運相關的4個ATP-binding cassette (ABC) transporter基因,并采用Reverse Transcription quantitative real-time PCR (RT-qPCR)技術分析在不同濃度的鋅離子脅迫下這4個ATP-binding cassette (ABC) transporter基因在轉錄水平上的差異表達情況。然后利用Vector NTI、cluster X、BLAST、ORF Finder等生物信息學軟件對各基因做進一步的生物信息學分析。結果表明:在鋅離子脅迫下,4個ABC transporter基因(AFE_2435、AFE_2436、AFE_2437、AFE_2438)的表達量均有所上調,說明這4個基因對于鋅離子的脅迫具有很強的敏感性。經生物信息學分析可知,基因AFE_2435和AFE_2436編碼位于細胞質膜上的透性酶,AFE_2437編碼位于細胞質膜上的ATP結合蛋白,AFE_2438編碼位于周質空間的鋅離子結合蛋白,這 4個基因共同組成了一個與鋅離子轉運有關的 ABC轉運子。這些結果都表明這4個 ABC transporter基因與DC菌中鋅離子的轉運有著密切的關系。
嗜酸氧化亞鐵硫桿菌;ABC transporter;RT-qPCR;鋅離子脅迫;生物信息學分析
嗜酸氧化亞鐵硫桿菌(Acidithiobacillus ferrooxidans)[1]是一種重要的浸礦菌種,在工業上經常被用作還原金、銀、銅、鈾等重金屬[2]。在這些菌種生活的環境中,通常都含有較高濃度的重金屬,因此,像大部分的浸礦菌一樣,Acidithiobacillus ferrooxidans(At. ferrooxidans)對很多重金屬都具有很高的耐受能力[3]。近年來,At. ferrooxidans對重金屬的這種高耐受能力越來越引起人們的關注[4]。盡管 At. ferrooxidans標準菌ATCC23270的全基因組序列已在2007年被全部測出,但是卻很少有基因[5?8]被證明與這種高抗性能的體現有直接關系。
鋅對于生物體來說是一種必須的微量元素,許多重要的功能蛋白和酶都需要鋅作為其結構或者是輔助因子[9?10]。但是,一旦體內的鋅離子濃度過高,將會對呼吸鏈產生抑制從而對細胞造成毒害作用[11?12]。因此,細胞需要調節其體內的鋅離子含量在一個合適的水平。近年來有研究報道At. ferrooxidans 對鋅離子具有很高的耐受能力,它可以在30 g/L的鋅離子環境中生存[13]。然而,什么機制使At. ferrooxidans具有如此高的抗鋅能力,至今仍不清楚。關于鋅離子的轉運機制,研究較多的是一種存在于大腸桿菌中的依賴于ATPase的 ABC轉運子—ZnuABC[14?15],它主要負責從外界環境中攝取鋅離子。通常來說,這種 ABC轉運子[14]主要由綁定在周質空間上的鋅離子結合蛋白ZnuA、兩個透性酶 ZnuB以及為此過程提供能量的ATP結合蛋白ZnuC組成。鋅離子的這種轉運機制在其他的菌種中也普遍存在[16?17],例如:肺炎鏈球菌(Streptococcus pneumoniae),格氏鏈球菌(Streptococcus gordonii),腸道沙門氏桿菌(Salmonella enterica)。
2007年 CHI等[18]對可能存在于 At. ferrooxidans ATCC2327周質空間上的蛋白進行了鑒定,推測AFE_2438所編碼的蛋白是一種位于周質空間上的鋅離子轉運蛋白,它的功能可能類似與ZnuABC中的鋅離子綁定蛋白ZnuA。而位于其下游的基因AFE_2437、AFE_2436、AFE_2435與其一起共同組成一個 ABC轉運子,完成鋅離子的轉運。盡管如此,至今仍沒有任何實驗數據證明這4個基因與At. ferrooxidan中的鋅離子轉運有關。
本文作者以這4個基因(AFE_2435、AFE_2436、AFE_2437、AFE_2438)作為研究對象,利用實時熒光定量 PCR技術[19]驗證在不同濃度的鋅離子刺激下它們在基因轉錄水平上差異表達情況,并通過生物信息學手段對這4個基因及其編碼的蛋白進行結構和功能的預測。
1 實驗
1.1 材料
1.1.1 菌種與培養基
At. ferrooxidans DC由中南大學生物冶金教育部重點實驗室從取自廣西大廠銅礦的酸性礦坑廢水中分離得到。
At. ferrooxidans DC生長于 9K基礎培養基[20],30 ℃,170 r/min 搖床無菌培養。能源物質為單質硫:10 g/L。
1.1.2 其他試劑
DNA提取試劑為 EZ-10 spin column genomic DNA isolution kit (BioBasic Inc.),DNA凝膠回收試劑盒為(E.Z.N.A.TMGel Extraction Kit.Promega),RNA提取試劑為Trizol(Invitrogen),RNA純化試劑盒為SV Total RNA Isolation System(Promega);RNA反轉錄試劑為SuperScriptTMⅡ反轉錄酶(Invitrogen).
1.2 實驗方法
1.2.1 菌種收集
前期預實驗表明At. ferrooxidans strain DC能夠耐受較高濃度的鋅離子,但是其生長繁殖情況會受到一定的抑制,本實驗中選用1、10和100 mmol/L Zn2+作為鋅離子刺激環境。首先將 At. ferrooxidans strain DC接種至不含鋅離子的標準培養基,培養至對數期時離心收集菌種(4 ℃, 10 min, 12 000 r/min),再將所收集的菌種等量接種于含有0、1、10和100 mmol/L Zn2+的9K培養基中,分別培養24 h后,再次離心收集菌種,馬上進行DNA和RNA提取步驟。
1.2.2 DNA提取及ZnuAf基因的克隆測序
細菌基因組提取使用EZ-10 spin column genomic DNA isolution kit。以提取后的DC基因組為模板擴增基因AFE_2435、AFE_2436、AFE_2437、AFE_2438。PCR擴增條件為:預變性:94 ℃,3 min;變性:94 ℃,45 s,退火:60 ℃,45 s,延伸:72 ℃,90 s,共30個循環;延伸:72 ℃,10 min。PCR產物經瓊脂糖凝膠電泳檢測后切膠,回收,送去上海生物工程公司測序。實驗中所用到的引物如表1所列。
1.2.3 RNA提取與cDNA的合成
采用Trizol一步法提取總RNA,用RNA純化試劑盒(Promega)純化粗RNA,用NanoDrop微量分光光度計(NanoDrop Technologies)檢測 RNA 的濃度和純度。取等量的RNA進行反轉錄,反轉錄采用Invitrogen公司 SuperScriptTMⅡ反轉錄酶和隨機引物,以總RNA中mRNA為模板,反轉錄合成cDNA,反轉錄后用NanoDrop 微量分光光度計測定每一個cDNA的濃度, 然后將cDNA樣品濃度均調至200 mg/L,于?20 ℃冷藏備用。
1.2.4 Real-time qPCR
1.2.4.1 標準樣品的制備
分別以目標基因及內參基因為樣品, 以cDNA為模板進行普通PCR 擴增,PCR產物經瓊脂糖凝膠電泳鑒定。然后將 PCR 產物進行 10倍梯度稀釋, 取10?3~10?7做標準品用于制備標準曲線, 做5個點。普通PCR程序如下:預變性:94 ℃,3 min;變性:94 ℃,30 s,退火:55 ℃,30 s,延伸:72 ℃,30 s, 共 35個循環;最后72 ℃補平5 min。
1.2.4.2 實時定量PCR
Real-time qPCR反應程序如下:預變性:95 ℃,3 min ;變性:95 ℃,30 s, 退火:59 ℃,20 s,延伸 72 ℃,20 s,共40個循環;55~95 ℃,10 s, 每循環一次溫度增加0.5 ℃,共80個循環。每組實驗設置3個平行,選用16S rRNA為內參基因[5],陰性對照不加任何模板。
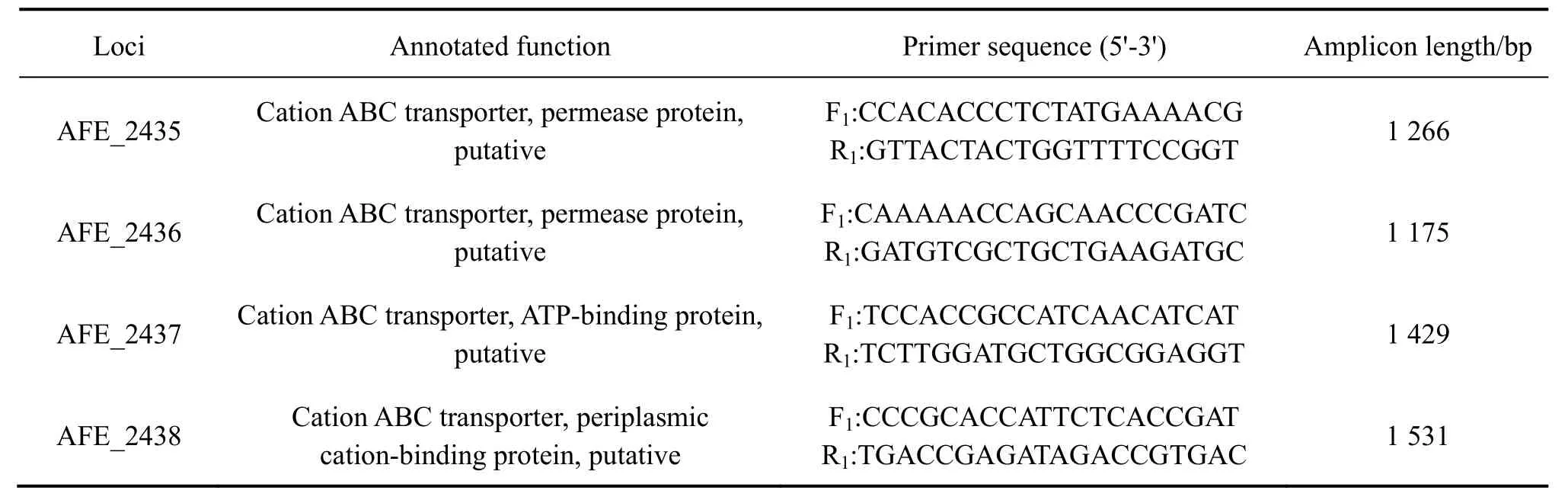
表1 常規PCR引物Table 1 Primers used in Taq PCR
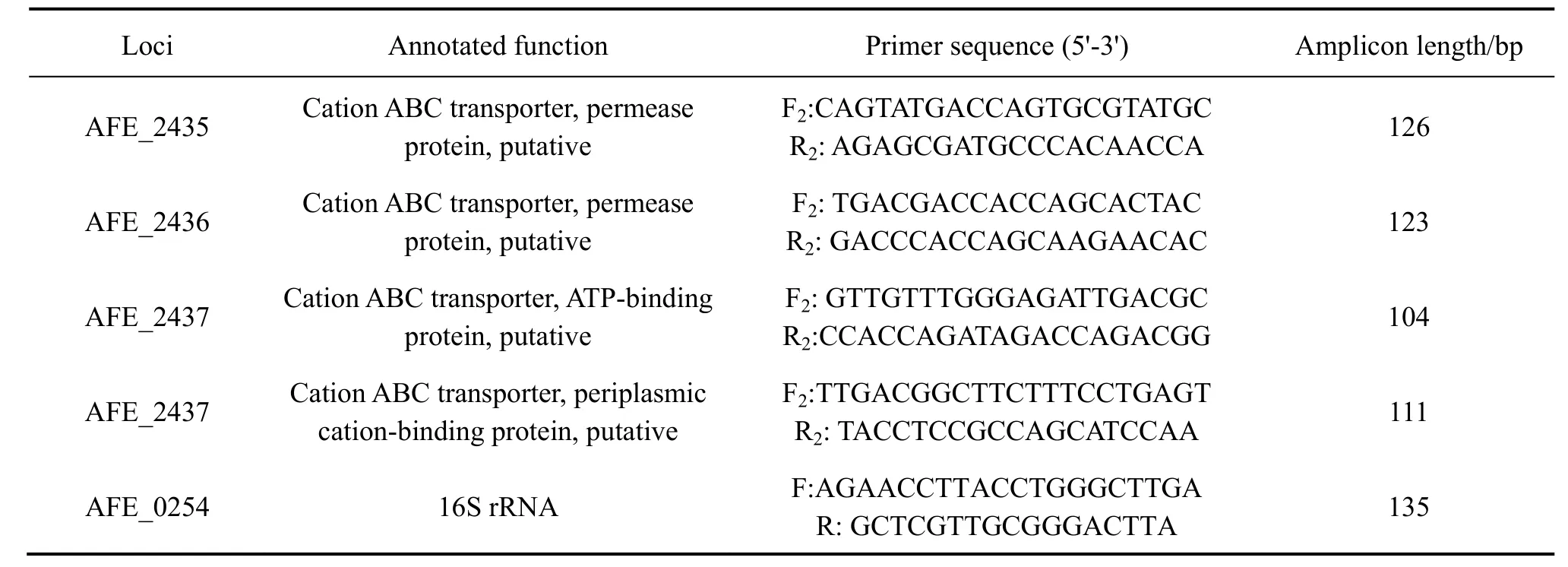
表2 RT-qPCR引物Table 2 Primers used in RT-qPCR
實驗結束后,用 2?△△Ct法[7,21]處理數據,即通過與對照組基因表達量的對比計算出每個基因的相對表達量,且每個基因都以16S rRNA作為內參基因進行了校正。實驗中所用到的引物如表2所列。
1.2.5 生物信息學分析
用Vector NTI (version 7.1)做一般序列操作。用cluster X 做序列比對。BLAST (http://www.ncbi.nih.gov/blast/Blast.cgi) 做相似性搜索。ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html) 尋找基因的開放閱讀框。ExPASy Proteomics Server (http://expasy. org/ cgi- bin/ pi_ tool) 進行蛋白質等電點以及相對分子量的計算。PSORTb v.3.0 (http://www.psort.org/psortb) 做蛋白亞細胞結構定位。TMHMM Server v.2.0 (http://www.cbs.dtu.dk// servive-s/TMHMM-2.0/)做蛋白質的跨膜分析。NCBI conserved domains (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) 找尋蛋白的保守區域。
2 結果
2.1 ZnuAf基因的克隆測序
以 At. ferrooxidans DC基因組為模板擴增得到AFE_2435、AFE_2436、 AFE_2437、AFE_2438 的基因片段,電泳檢測結果如圖1所示,擴增產物長度與目標基因長度基本相同,且均無雜帶,引物特異性較好。4個基因的測序結果經BLAST進行序列比對,結果與嗜酸氧化亞鐵硫桿菌標準菌株ATCC 23270中的這4個基因的序列完全相同。

圖1 目的基因 AFE_2435、AFE_2436、AFE_2437、AFE_2438的PCR產物電泳圖Fig. 1 Electrophoresis analyses of PCR products of AFE_2435, AFE_2436, AFE_2437 and AFE_2438
2.2 不同濃度Zn2+刺激下相關基因的差異表達
實時熒光定量PCR后,溶解曲線峰帶單一,沒有雜峰出現,說明特異性較好;標準曲線R2值均接近于1,說明Ct值與其起始拷貝數的對數值之間的關系性好; 內參基因16S rRNA在不同環境中的表達量均較為衡定,無顯著性差異。經內參基因校正,各基因在轉錄水平上的相對表達量如圖2所示。在不同濃度的鋅離子刺激下,4個基因(AFE_2435、AFE_2436、AFE_2437、AFE_2438 ) 的表達量均有所上調,且隨著鋅離子濃度的增加,各基因上調的倍數也有所增加。AFE_2435和AFE_2436上調最為明顯,1 mmol/L Zn2+刺激時,AFE_2435上調了16倍,AFE_2436上調了33倍,而此時AFE_2438僅上調了9倍,AFE_2437僅上調了3倍。當鋅離子濃度增加到10 mmol/L時,AFE_2435上調的倍數高達230倍,AFE_2436上調的倍數為270倍,AFE_2438上調的倍數為77倍,雖然此時AFE_2437的表達量也有所增加,但是與其他3個基因相比較,AFE_2437的上調幅度較小,當鋅離子濃度為10 mmol/L時僅上調了5倍。當鋅離子濃度為100 mmol/L時,4個基因的表達量與未加鋅離子刺激的基因表達量相比較,均呈現出明顯的上調趨勢,AFE_2435上調了 442倍, AFE_2436上調了 512倍,AFE_2437上調了22倍, AFE_2438上調了321倍。
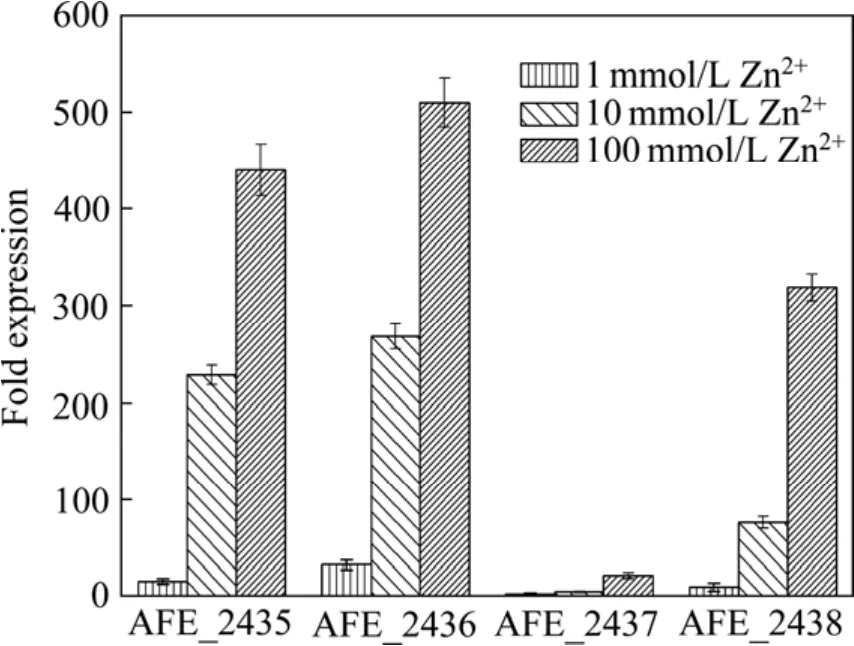
圖2 不同鋅離子刺激條件下 AFE_2435、AFE_2436、AFE_2437、AFE_2438的差異表達Fig. 2 Differential expressions of genes (AFE_2435,AFE_2436, AFE_2437 and AFE_2438) under different Zn2+concentrations
2.3 生物信息學分析
本研究對實驗中涉及到的 4個基因(AFE_2435、AFE_2436、AFE_2437、AFE_2438) 以及其編碼的蛋白進行了相關的生物信息學分析。在此將它們所編碼的蛋白分別命名為:ZnuB1Af、ZnuB2Af、ZnuCAf和ZnuAAf。通過對 4個基因進行開放閱讀框的找尋以及等電點和分子量的計算可知:基因AFE_2435的開放閱讀框的長810 bp;它所編碼的蛋白ZnuB1Af的相對分子質量為 28 572.27 Da,等電點為 10.1。基因AFE_2436含有一個長為891 bp開放閱讀框;它所編碼的蛋白ZnuB2Af的相對分子質量為30 704.72 Da,等電點為9.04。基因AFE_2437的開放閱讀框長825 bp,ZnuCAf的相對分子質量為 29 068.60 Da,等電點為6.66。而基因 AFE_2438的開放閱讀框的長度為 513 bp,其編碼的蛋白ZnuAAf的相對分子質量為31 739.21 Da, 等電點為8.51。
為了更進一步了解這4個蛋白所行使的功能,還對 ZnuB1Af等 4個蛋白進行了亞細胞結構定位、跨膜結構域分析以及保守區域的分析。結果表明:ZnuB1Af與ZnuB2Af均位于質膜上,其中ZnuB1Af有6次跨膜,ZnuB2Af有 7次跨膜,保守區域分析表明 ZnuB1Af與ZnuB2Af含有相同的保守區域cd06550,該保守區域是TM_ABC_iron-siderophores_like super family所特有的; ZnuCAf也位于細胞質膜上,但是它并不存在跨膜結構域,經比對發現 ZnuCAf的保守區域與 P-loop NTPase surper family家族中的蛋白高度相似,而且ZnuCAf還具有與該家族蛋白相同的保守基序(-LSGGQ);ZnuAAf是4個蛋白中唯一一個位于周質空間上的蛋白,它也不存在跨膜結構域,它的保守區域與TroA-like family中的保守區cd01020高度相似。
3 討論與分析
本實驗中,對At. ferrooxidans DC中4個與鋅離子轉運相關的基因(AFE_2435、AFE_2436、AFE_2437、AFE_2438)進行了克隆、測序,并應用實時熒光定量PCR(RT-qPCR)[22?24]技術來檢測不同濃度鋅離子刺激下這4個基因的相對表達量。實驗中所選用的內參基因為16S rRNA。雖然近年來有研究認為,在某些情況下以16S rRNA作為內參基因校正基因的相對表達量時可能存在誤差[25],但是在本實驗中16S rRNA在不同鋅離子環境中的表達量均相對穩定,因此可以作為內參基因。RT-qPCR結果表明,對于鋅離子的脅迫,這4個基因都具有很強的敏感性,在較低濃度(1 mmol/L)刺激時, AFE_2435和AFE_2436的反應較大,分別上調了16倍和33倍數;隨著鋅離子濃度的升高,其它兩個基因的表達量也出現了明顯的上調趨勢,當鋅離子刺激濃度達到100 mmol/L時,AFE_2437上調了22倍,AFE_2438上調了321倍,而此時AFE_2435和AFE_2436的上調倍數分別達到了442倍和512倍。此實驗數據表明,At. ferrooxidans DC中的這4個基因(AFE_2435, AFE_2436, AFE_2437, AFE_2438) 對于鋅離子的脅迫非常敏感。這4個基因可能與其能夠耐受較高濃度的鋅離子有著密切的關系。
生物信息學的分析結果表明:基因AFE_2435和AFE_2436所編碼的蛋白ZnuB1Af與ZnuB2Af與位于質膜上的鐵載體蛋白有著相同的保守區域,這個家族中的蛋白通常是作為ABC轉運子[26]的一個組件而存在。ZnuCAf的保守區域屬于P-loop NTPase surper family,該家族中的蛋白含有ATP或者GTP的結合位點,它們與iron-siderophores uptake family擁有共同的祖先,這兩個家族中的蛋白都與鋅離子、錳離子、鐵離子的轉運密切相關。ZnuAAf屬于TroA-like family,這個家族中的蛋白在 ABC轉運子轉運金屬離子的過程中擔當金屬離子的受體。綜上所述,可以推斷在 At.ferrooxidans DC轉運鋅離子的過程中,ZnuB1Af、ZnuB2Af、 ZnuCAf、 ZnuAAf共同組成了一個轉運 Zn2+的ABC轉運子,其中ZnuAAf主要負責在周質空間中綁定鋅離子,ZnuB1Af和ZnuB2Af作為透性酶負責鋅離子向細胞內的運輸,ZnuCAf作為ATP結合蛋白為此過程提供能量。
4 結論
1) 首次鑒定了At. ferrooxidans DC中與Zn2+轉運相關的4個ATP-binding cassette (ABC) transporter 基因。
2) 在不同濃度鋅離子刺激下,At. ferrooxidans DC中的這 4個 ABC transporter 基因(AFE_2435,AFE_2436, AFE_2437, AFE_2438)的相對表達量均呈上調趨勢,說明這4個ABC transporter 基因對于鋅離子的脅迫非常敏感。
3) 經生物信息學分析可知基因 AFE_2435和AFE_2436所編碼的蛋白是位于細胞質膜上的透性酶,基因AFE_2437編碼的蛋白也位于細胞質膜上,是一種ATP結合蛋白,而基因AFE_2438編碼位于周質空間的鋅離子結合蛋白,這4個基因共同組成了一個與鋅離子轉運有關的ABC轉運子。
4) 推斷 At. ferrooxidans DC中的這 4個 ABC transporter 基因(AFE_2435、AFE_2436、AFE_2437、AFE_2438)對于其能夠耐受較高濃度的鋅離子刺激有著密切的關系。
REFERENCES
[1] INGLEDEW W J. Thiobacillus ferrooxidans: The bioenergetics of an acidophilic chemolithotroph[J]. Biochim Biophys Acta,1982, 683(2): 89?117.
[2] RAWLINGS D E. Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates[J]. Microbial Cell Factories,2005, 4: 13.
[3] DOPSON M, BAKER-AUSTIN C, KOPPINEEDI P R, BOND P L. Growth in sulfidic mineral environments: Metal resistance mechanisms in acidophilic microorganisms[J]. Microbiology,2003, 149: 1959?1970.
[4] RAWLINGS D E, KUSANO T. Molecular genetics of Thiobacillus ferrooxidans[J]. Microbiology and Molecular Biology Reviews, 1994, 58(1): 39?55.
[5] NAVARRO C A, ORELLANA L H, MAURIACA C, JEREZ C A. Transcriptional and functional studies of Acidithiobacillus ferrooxidans genes related to survival in the presence of copper[J]. Applied Environmental Microbiology, 2009, 75:6102?6109.
[6] LUO Y J, LIU Y D, ZHANG C G, LUO H L, GUAN H, LIAO H H, QIU G Z, LIU X D. Insights into two high homogenous genes involved in copper homeostasis in Acidithiobacillus ferrooxidans[J]. Current Micobiology, 2008, 57: 274?280.
[7] FERRAZ L F C, VERDE L C L, REIS F C, ALEXANDRINOl F,FELI′CIO A P, NOVO M T M, GARCIA O Jr, OTTOBONI L M M. Gene expression modulation by chalcopyrite and bornite in Acidithiobacillus ferrooxidans[J]. Archives of Microbiology,2010, 192: 513?540.
[8] PAULINO L C, de MELLO M P, OTTOBONI L M M.Differential gene expression in response to copper in Acidithiobacillus ferrooxidans analyzed by RNA arbitrarily primed polymerase chain reaction[J]. Electrophoresis, 2002, 23:520?527.
[9] VALLEE B L, AULD D S. Zinc coordination, function, and structure of zinc enzymes and other proteins[J]. Biochemistry,1990, 29(24): 5647?5659.
[10] PATZER S I, HANTKE K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli[J].Molecular Microbiology, 1998, 28(6): 1199?1210.
[11] KASAHARA M, ANRAKU Y. Succinate- and NADH oxidase systems of Escherichia coli membrane vesicles: Mechanism of selective inhibition of the systems by zinc ions[J]. The journal of Biochemistry, 1974, 76(5): 967?976.
[12] BEARD S J, HUGHES M N, POOLE R K. Inhibition of the cytochrome bd-terminated NADH oxidase system in Escherichia coli K-12 by divalent metal cations[J]. FEMS Microbiol Letters,1995, 131(2): 205?210.
[13] CABRERA G, GO′MEZ J M, CANTERO D. Influence of heavy metals on growth and ferrous sulphate oxidation by Acidithiobacillus ferrooxidans in pure and mixed cultures[J].Process Biochemisty, 2005, 40: 2683?2697.
[14] PATZER S I, HANTKE K. The zinc-responsive regulator Zur and its control of the znu gene cluster encoding the ZnuABC zinc uptake system in Escherichia coli[J]. The journal of Biological Chemistry, 2000, 275: 24321?24332.
[15] GRASS G, WONG M D, ROSEN B P, SMITH R L, RENSING C. ZupT is a Zn(Ⅱ) uptake system in Escherichia coli[J]. Journal of Bacteriology, 2002, 184: 864?866.
[16] NIES D H. Microbial heavy-metal resistance[J]. Applied Microbiology and Biotechnology, 1999, 51: 730?750.
[17] CAMPOY S, JARA M, BUSQUETS N, PE′REZ de ROZAS A M, BADIOLA I, BARBE J. Role of the high-affinity zinc uptake znuABC system in Salmonella enterica serovar typhimurium virulence[J]. Infection and Immunity, 2002, 70: 4721?4725.
[18] CHI A, VALENZUELA L, BEARD S, MACKEY A J,SHABANOWITZ J, HUNT D F, JEREZ C A. Periplasmic proteins of the extremophile Acidithiobacillus ferrooxidans[J].Molecular and Cellular Proteomics, 2007, 6(12): 2239?2251.
[19] ZAMMINT C M, MUTCH L A, WATLING H R, WATKIN E L J. Evaluation of quantitative real-time polymerase chain reaction for enumeration of biomining microorganisms in culture[J].Hydrometallurgy, 2008, 94: 185?189.
[20] WU Xue-ling, DING Jian-nan, GAO Jian, LIU Xing-xing, QIU Guan-zhou. Isolation and identification of metal-resistant iron-oxidizing bacteria[J]. Minerals and Metallurgical Processing,2007, 24: 57?60.
[21] XU Ai-ling, XIA Jin-lan, LIU Ke-ke, LI Li, YU Yang, NIE Zhen-yuan. Real-time PCR analysis of metabolic pathway of PHB in acidiphilium cryptum DX1-1[J]. Journal of Microbiology and Biotechnology, 2010, 20: 71?77.
[22] 劉小榮,張 笠, 王勇平. 實時熒光定量 PCR技術的理論研究及其醫學應用[J]. 中國組織工程研究與臨床康復, 2010,14(2): 329?332.LIU Xiao-rong, ZHANG Li, WANG Yong-ping. Theory study and medical application of real-time quantitative polymerase chain reaction[J]. Journal of Clinical Rehabilitative Tissue Engineering Research, 2010, 14(2): 329?332.
[23] 徐小剛, 劉雅婷. 實時熒光定量PCR在植物病害中的應用[J].中國農學通報, 2009, 25(7): 52?56.XU Xiao-gang, LIU Ya-ting. Application of real-time fluorescence quantitative PCR in plant disease[J]. Chinese Agricultural Science Bulletin, 2009, 25(7): 52?56.
[24] SCHMITTGEN T D, ZAKRAJSEK B A, MILLS A G, GORN V,SINGER M J, REED M W. Quantitative reverse transcription-polymerase chain reaction to study mRNA decay:Comparison of endpoint and real-time methods[J]. Analytical Biochemistry, 2000, 285: 194?204.
[25] NIETO P A, COVARRUBIAS P C, JEDLICKIL E, HOLMES DS, QUATRINI R. Selection and evaluation of reference genes for improved interrogation of microbial transcriptomes: Case study with the extremophile Acidithiobacillus ferrooxidans[J]. BMC Molecular Biology, 2009, 10:63. doi:10.1186/1471-2199/10/63.[26] LOCHER K P. Structure and mechanism of ATP-binding cassette transporters[J]. Philosophical Transaction of the Royal Society, 2009, 364: 239?245.
Identification and analysis of zinc transport genes in Acidithiobacillus ferrooxidans
HOU Dong-mei1, MIAO Bo1, WANG Yang-yang1, ZHANG Yun-jing1, LIU Dai-gang1, WU Xue-ling2
(1. School of Minerals Processing and Bioengineering, Changsha 410083, China;2. Key Laboratory of Biometallurgy, Ministry of Education, Central South University, Changsha 410083, China)
In this study, four ATP-binding cassette (ABC) transporter genes of Acidithiobacillus ferrooxidans DC (At.ferrooxidans DC) were identified, and differential transcription of these genes during zinc ion stress were investigated by Reverse Transcription quantitative real-time PCR (RT-qPCR). And then, the genes involved in zinc ion transport were analyzed by bioinformatics software. The results show that the expressions of the four ABC transporter genes are increased differently under zinc ion stress, indicating that these genes are sensitive to zinc levels. Bioinformatics analysis shows that the proteins encode by AFE_2435, AFE_2436 are predicted to be permease proteins, whereas the protein encode by AFE_2437 is a putative ATP-bindind protein and AFE_2438 encoded a putative periplasmic cation-binding protein. The four genes together form an ABC transporter for zinc ion transport. These results strongly suggest that the four ABC transporter genes might be directly involved in zinc transport in Acidithiobacillus ferrooxidans DC.
Acidithiobacillus ferrooxidans; ABC transporter; RT-qPCR; zinc ion stress; bioinformatics analysis
Q819
A
1004-0609(2012)05-1497-06
國家重點基礎研究發展計劃資助項目(2010CB630901)
2011-02-17;
2011-05-09
吳學玲,副教授,博士;電話:0731-88836944;傳真:0731-88710804;E-mail: xueling0714@yahoo.com.cn
(編輯 何學鋒)
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
