3個毛白楊病程相關蛋白基因的克隆及表達1)
2012-09-18 11:11:06張志毅劉文鳳安新民林善枝
東北林業大學學報 2012年6期
關鍵詞:分析
雷 楊 張志毅 劉文鳳 王 興 安新民 林善枝
(林木育種國家工程實驗室(北京林業大學),北京,100083)
鄭會全
(廣東省林業科學研究院)
植物在自然界往往受到各種生物與非生物脅迫的不利影響,因此經過漫長演化過程,產生了一系列復雜的防御機制。病程相關蛋白(pathogenesis related proteins,PRs/PRP)是植物在各種病原物的侵染及所引起的相關脅迫(病原菌、線蟲、昆蟲、食草動物等生物脅迫),以及模擬病原物侵害效應或類似脅迫的化學試劑(水楊酸(SA)、乙烯(ET)、脫落酸(ABA)、茉莉酸(JA)等激素)的施用下誘導產生并積累的一類蛋白質的總稱,可作為下游防衛基因,是植物防衛反應體系的重要組成部分[1-2]。PRP最初由Van Loon等[2]從被煙草花葉病毒侵染的煙草葉片中分離獲得,隨著研究的深入,人們發現PRs廣泛存在于不同植物中,并逐漸形成了以蛋白序列的同源性、等電點、電泳遷移率(相對分子質量)、植物來源、生理活性以及血清學關系為標準的分類體系,進而將其分為17個家族[3-4]。其中PR-1雖具有抗真菌活性,但功能和作用機制至今未知[5];PR-2具有 β-1,3-葡聚糖酶活性,PR-3、PR-4、PR-8 和PR-11具有幾丁質酶活性,可抵御真菌;PR-5為類奇甜蛋白;PR-6為蛋白酶抑制劑,能夠防御昆蟲和線蟲;PR-7、PR-9、PR-10 分別為蛋白酶、過氧化物酶和核酶;PR-12~PR-16分別由防御素、硫素、轉脂蛋白、草酸氧化酶和類草酸氧化酶組成;PR-17含有類似鋅蛋白酶活性位點的序列,但具體功能未知。
盡管目前已從眾多物種中分離獲得 PRs,但PRs受誘導表達在物種間差異很大,并非所有的PR家族均存在于每一物種中;而且以往研究多集中于擬南芥、煙草、水稻、大麥、玉米、苜蓿等草本植物。楊樹作為木本模式植物[6]和重要的人工栽培林樹種,其抗病研究一直是林木抗逆育種的熱點。毛白楊(Populus tomentosa Carr.)是我國特有的白楊派鄉土樹種,具有分布廣、速生、抗逆性強、材質優良等特性[7]。因此,本文以毛白楊為試材,經過SA和Me-JA誘導處理,分離獲得多個編碼病程相關蛋白的基因序列,并運用生物信息學和實時熒光定量PCR技術,對基因結構和誘導表達水平進行比較分析,從中探討木本植物病程相關蛋白的功能及抗病信號傳導網絡對防衛反應基因表達的調控機理。
1 材料與方法
用于誘導處理的毛白楊組培苗保存于北京林業大學林木花卉遺傳育種教育部重點實驗室和林木育種國家工程實驗室。將組培苗移栽至土壤中2個月后,選取生長狀態一致的植株作為試驗材料。
材料處理:以移栽2個月后的毛白楊植株為試材,向每株葉片完全展開的區域(2~4葉片間)分別噴灑10 mL外源SA溶液(5 mmol/L)和MeJA溶液(200 μmol/L),以噴灑相同體積的雙蒸水(ddH2O)為對照。各種處理后的材料用透明袋密封,在處理6、12、24和48 h后,分別采集處理過的葉片。
總DNA與總RNA的提取及第一鏈cDNA的合成:對采集的經處理葉片分別提取總 DNA和總RNA。總 DNA的提取采用 Tiangen植物基因組DNA 提取試劑盒(Tiangen Biotech Co.,Ltd.,Beijing);總 RNA的提取依照 SV Total RNA Isolaton System 試劑盒(Promega Biotech Co.,Ltd.,USA)的方法進行。以總RNA為模板,采用Reverse Transcription System(Promega Biotech Co.,Ltd.,USA)合成第一鏈cDNA。
RT-PCR引物設計和擴增:通過分析楊樹基因組數據庫,獲得病程相關蛋白基因PR-1、PR-5、PR-10的電子克隆序列,據此設計引物,如表1所示。PCR 擴增體系為 25 μL(其中 cDNA 2 μL,MgCl21.5 mmol/L,dNTPs 0.8 mmol/L,上下游引物各 0.4 μmol/L,Taq酶1.25 U)。反應程序為:94℃預變性4 min;94℃變性30 s、58℃退火30 s、72℃延伸40 s,循環35次;72℃延伸10 min。反應產物通過1.1%瓊脂糖凝膠電泳檢測并進行分離回收和純化。
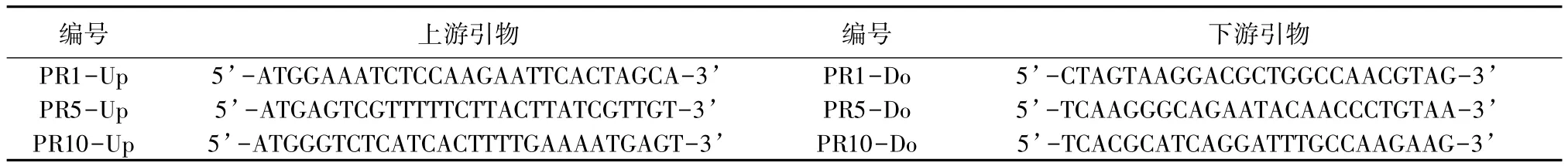
表1 用于RT-PCR反應的引物序列
克隆與測序:將PCR回收產物與pGEM-T載體連接,轉化Top10大腸桿菌后,利用菌落PCR擴增輔以藍白斑篩選鑒定陽性克隆。將陽性克隆單菌落搖菌,送GENEWIZ公司測序。
生物信息學分析:測序結果先通過DNASTAR Lasergene v7.1推測出蛋白序列,再經NCBI(http://www.ncbi.nlm.nih.gov/)和 SMART(http://smart.embl-heidelberg.de/)分析同源性和蛋白序列保守結構域,SWISS-MODEL(http://swissmodel.expasy.org/)模擬蛋白三維結構[8-10]。使用 ClustalX2.0、BioEdit7.01 和 MEGA4.1,采用鄰接法(Neighbor-Joining,N-J)構建系統發育樹,并使用1000次重復的自展檢驗評估系統發育樹拓撲結構的穩定性。此外,通過 CBS(http://www.cbs.dtu.dk/index.shtml)在線分析信號肽、磷酸化位點等信息。
實時熒光定量RT-PCR分析:將不同處理的毛白楊葉片總RNA分別合成第一鏈cDNA后,通過DNA Engine Opticon 2 Real-Time Cycler熒光定量分析儀使用SYBR Premix ExTaq試劑盒(TaKaRa Biotechnology(Dalian)Co.,Ltd.,Dalian),采用 SYBR Green I相對定量法對病程相關蛋白基因的表達進行分析。選用ACTIN作為內參基因,引物序列參照Regier等[11];對應不同的病程相關蛋白基因的qRTPCR引物的設計,參照Rinaldi等[12],并由 Invitrogen公司合成,序列如表2所示。

表2 用于qRT-PCR反應的引物序列
2 結果與分析
2.1 毛白楊病程相關蛋白基因的克隆與分析
2.1.1 PtPR-1 基因序列的分析
毛白楊PR-1基因(HM589199)無內含子(圖1),讀碼框為486 bp,可編碼一個161aa的前體蛋白,包含136aa的成熟肽和一段25aa的信號肽。去除信號肽的成熟PtPR-1蛋白,包含一個SCP結構域,分子量為14.6 KD,等電點為8.24,為堿性蛋白。經預測后分析,PtPR-1蛋白序列中無糖基化位點,而其磷酸化位點可能位于3處絲氨酸(Ser28、128、150),2處蘇氨酸(Thr66、121)和 6處酪氨酸(Tyr32、57、78、105、107、120)(圖 2)。

圖1 毛白楊病程相關蛋白基因PR-1、PR-5、PR-10的PCR擴增

圖2 PtPR-1、PtPR-5、PtPR-10 基因序列分析
多重序列比對結果顯示,雖然信號肽序列差異顯著,但毛白楊PR-1前體蛋白序列的相似度與其他物種相比仍保持在很高的水平,分別為擬南芥(Arabidopsis thaliana,P33154.1)63%、馬鈴薯(Solanum phureja,Q08697.1)53%、西洋接骨木(Sambucus nigra,Q41359.1)62%、煙草(Nicotiana tabacum,P07053.1)60%(圖3)。系統發育樹分析結果表明,多物種間PR-1家族同源性很高(圖4)。另外,PtPR-1前體蛋白中的信號肽為α螺旋結構,而模擬的成熟蛋白結構含有3個α螺旋、4個β折疊結構、6個半胱氨酸形成3個二硫鍵(圖4)。

圖3 PtPR-1、PtPR-5、PtPR-10 氨基酸序列多重比對
2.1.2 PtPR-5 基因序列的分析
毛白楊PR-5基因(HM589200)無內含子(圖1),讀碼框為678 bp,可編碼一個225aa的前體蛋白,包含200aa的成熟肽和一段25aa的信號肽;而去除信號肽的成熟PtPR-5蛋白(分子量21.7KD、等電點6.21),為酸性蛋白,包含一個THN結構域,其核心結構為 G-x-[GF]-x-C-x-T-[GA]-D-C-x-(1,2)-G-x-(2,3)-C[13-14]。經預測分析,PtPR-5蛋白序列中可能的磷酸化位點位于3處絲氨酸(Ser5、123、161),2 處蘇氨酸(Thr73、105)和 2 處酪氨酸(Tyr176、200)(圖2)。與其他物種PR-5家族序列多重比對后發現,除信號肽序列顯著差異外,毛白楊PR-5前體蛋白序列與擬南芥(Arabidopsis thaliana,NP_177641.1)、沙梨 (Pyrus pyrifolia,BAC78212.1)、地中海柏木(Cupressus sempervirens,AAR21075.1)、煙草滲透素(Nicotiana tabacum osmotin,P14170.2)以及煙草 PR-5d(Nicotiana tabacum PR-5d,P25871.1)的相似性分別為 45%、39%、48%、67%和62%(圖3)。另外,PtPR-5前體蛋白中的信號肽為α螺旋結構,而模擬的成熟蛋白包含17處卷曲(幾乎全部暴露在蛋白分子外側)、6個β折疊結構近似排列成桶狀(全部位于蛋白內部)、且有17個半胱氨酸形成二硫鍵(圖4)。

圖4 PtPR-1、PtPR-5、PtPR-10蛋白序列的系統發育樹及其三維結構模擬圖
2.1.3 PtPR-10 基因序列的分析
毛白楊PR-10基因組序列大小為574 bp包含了一個91-bp的內含子序列(圖1),開放閱讀框為483 bp(HM589201),可編碼一個分子量17.9 KD、等電點4.82的160aa蛋白,為酸性蛋白,包含一個Bet_v_1結構域。經預測分析后發現,PtPR-10蛋白序列中可能的磷酸化位點位于3處絲氨酸(Ser38、41、85),4 處蘇氨酸(Thr53、58、109、121)和 4 處酪氨酸(Tyr82、102、122、139)(圖 2)。多重序列比對分析結果表明,毛白楊PR-10蛋白序列相似性一般,例如歐洲白樺(Betula pendula,CAB94733.1)46%、剛毛檉柳(Tamarix hispida,ACK38253.1)40%、加州山松(Pinus monticola,AAL49994.1)28%、蘋果(Malus domestica,AAS00053.1)32%、刺茄(Solanum virginianum,AAU00066.1)41%(圖 4)。另外,蛋白模擬結構如圖4所示,PtPR-10主體結構為7個β折疊、2條長短不一的α螺旋和1個雙硫鍵。
2.2 PtPR-1、PtPR-5 和 PtPR-10 的誘導表達分析
為了研究SA、MeJA誘導處理對毛白楊的轉錄水平表達變化的影響,本實驗采用實時熒光定量PCR的方法分別對誘導處理不同時間后的PtPR-1、PtPR-5和PtPR-10的基因表達量進行檢測。結果如圖5所示,PtPR-1在SA誘導下初始表達水平較低,24 h后表達量顯著增加,48 h后已迅速升高約為初始水平200倍。另外,在MeJA處理下,PtPR-1呈先高后低再劇增的特征,初期PtPR-1表達量開始升高(于6 h后達到小高峰、約為初始水平100倍),12 h后降至接近初始水平,但在隨后誘導24至48 h期間其表達水平劇增至約為初始水平的400倍。
與PtPR-1相比,PtPR-5無論在SA還是MeJA誘導下表達量均相對較低(圖5)。PtPR-5的表達受SA誘導影響,呈先高后低再恢復初始水平的波動特點;在MeJA影響下,PtPR-5表達先降低再恢復初始水平。
PtPR-10的誘導表達模式與 PtPR-1、PtPR-5均有所不同(圖5)。在SA作用下,PtPR-10表達量有所提高,6 h即達到最高值,隨后呈緩慢遞減趨勢;而在MeJA誘導下,PtPR-10表達水平不斷升高,這與PtPR-1受SA誘導表達模式相似,但增長曲線更加平緩、且增長值也較小。
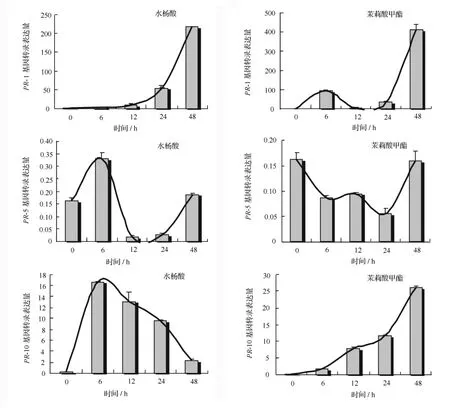
圖5 水楊酶、抹莉酸P酯誘導處理對毛白楊PR-1、PR-5和PR-10的轉錄水平變化的影響
3 討論
植物受病原菌侵染后,局部受感染部分細胞迅速凋亡,進而激發植物體對后續病原物產生廣譜、持久和系統的抗性,即所謂的系統獲得性抗性(Systemic acquired resistance,SAR)[15]。SA 是植物體內含量較低的一種內源酚類物質,目前已有大量實驗證實,SA是誘導SAR的關鍵信號分子,可抑制過氧化氫酶(CAT)和過氧化物酶的活性而造成H2O2的累積[16-18]。H2O2一方面作可為強氧化劑而能直接抵御病原菌的侵害,但對植物體自身也造成較大的毒害作用;另一方面,H2O2與活性氧的積累對防衛反應基因的激活可起著第二信使的作用。另外,JA及其揮發性衍生物MeJA不僅影響植物體的生長發育,而且也是種創傷誘導的內源信號分子,可啟動相關防衛基因的表達。目前已有報道,植物主要依靠SA、JA等多種信號通路間的“crosstalk”互作機制,調控包括病程相關蛋白基因在內的大量防衛反應相關基因的表達,在植物基礎免疫系統中發揮這重要作用[19]。
煙草PR-1蛋白相關研究證實,PR-1家族分為堿性蛋白和酸性蛋白兩種形態,其中酸性PR-1蛋白分布在細胞間隙,受SA信號通路誘導表達;而堿性PR-1蛋白則分布在液泡中,受JA/ET信號通路誘導[20-24]。在本研究中,對毛白楊 PtPR-1 的 N端信號肽、C端糖基化位點和分子量等進行分析,結果均顯示為典型的酸性PR-1蛋白特征,但等電點分析顯示為偏堿性蛋白。Ward等[25]研究后發現,同時施用SA和MeJA的混合物,酸性PR-1的誘導表達量明顯高于單獨施用SA時的水平。由此可推測,毛白楊PtPR-1可能兼具PR-1家族酸性蛋白和堿性蛋白的特點,在SA和MeJA處理下均受誘導表達。目前,眾多研究證實,NPR1(nonexpressor of pathogenesis-related genes 1)是SAR信號轉導途徑中作用于SA下游的關鍵性調控因子,而SA可誘導細胞內氧化還原態發生變化,無活性的NPR1寡聚體可被還原激活為NPR1單體,并轉運至細胞核內與結合在PR基因啟動子區的轉錄因子TGA互作激活 PR-1 基因[26-30]。同時,Petersen 等[31]通過對擬南芥MAPKs通路的研究后發現,AtMPK4蛋白元件既可作為負調節因子下調植物體SA水平而抑制SAR,又可作為正調節因子激活JA響應基因的表達。本實驗中施用高濃度外源SA后發現,毛白楊體內SA水平提高,推測可能是在NPR1介導下促進了毛白楊PtPR-1的表達,而毛白楊體內SA水平增高可造成H2O2積累及氧迸發,可能激發MAPKs通路中SA的負調控因子下調內源SA的水平,進而減弱了PtPR-1的表達,但同時也激活了JA響應基因的表達。Creelman等[32]實驗發現,SA-JA 在低濃度時存在協同作用,并且SA對JA的誘導作用強于JA對SA的誘導作用;Repka等[33]研究也證實SA處理可促進JA生物合成的一個關鍵酶AOS(丙二烯氧化物合酶)基因的表達;Rao等[34]發現在臭氧誘導的細胞死亡過程中,JA能抑制SA的生物合成和氧的迸發,從而減弱由SA引起的細胞死亡。據此推測,正是在減弱和增強雙重作用下,本實驗中毛白楊PtPR-1基因表達量增長緩慢,但隨著SA誘導時間的延長,初期毛白楊對內源SA的抑制作用趨于相對飽和,而持續高濃度的SA對PtPR-1誘導作用逐漸增大,同時內源JA積累量提高或許也有助于促進PtPR-1表達量的迅速放大。
另外,外源MeJA(50 μmol/L)能誘導大部分植株或懸浮培養細胞中合成SA的關鍵酶PAL的mRNA 的合成和酶活性的提高[33];但 Leon-Reyes等[35-36]對擬南芥 npr1突變體進行比較研究后發現,細胞質中的NPR1單體通過作用于JA合成途徑的下游,在SA-JA“crosstalk”中發揮拮抗作用而抑制JA響應基因的表達。因此,MeJA脅迫處理后的毛白楊植株,在促進PtPR-1表達同時,也可通過誘導SA合成酶基因PAL的表達而提高體內SA水平,進而發揮拮抗作用;但由于MeJA對SA的誘導作用相對較弱,而SA積累緩慢致使在處理初期對MeJA信號通路的拮抗作用并不顯著[32],這可能正是本研究中PtPR-1表達量保持增長趨勢的原因所在。目前,Mao等[37]研究顯示,擬南芥 WRKY62轉錄因子作用于細胞質中NPR1的下游并且負調控JA通路應答基因的表達,說明其很可能參與NPR1介導的SA對JA信號通路的拮抗過程。由此推測認為,初期隨著毛白楊體內SA積累量的提高,可能使得WRKY轉錄因子及NPR1介導拮抗作用的增強而降低PtPR-1表達量;但由于外源高濃度MeJA所造成的持續誘導效應強于內源SA對MeJA的拮抗,最終可促進PtPR-1表達量迅速恢復提高。
PtPR-5含有一個thaumatin保守結構域(THN結構域),與奇甜蛋白具有較高同源性,屬于PR-5蛋白家族中類甜蛋白(Thaumatin-Like Protein,TLP)。蛋白結構分析顯示,PtPR-5至少具有8個二硫鍵,說明其穩定性相對較高,可對蛋白酶具有很強的耐受性。另外,有些研究證實,PR-5能夠直接插入真菌細胞膜形成穿孔而造成水涌入細胞引發滲透性破裂,進而起到裂解孢子、抑制孢子萌發和菌絲生長的作用,表現出抗真菌活性[38,14]。但本研究發現,PtPR-5受SA和MeJA誘導作用并不顯著,這說明并非所有PR-5蛋白均能發揮活性,因此,PtPR-5蛋白在響應多種脅迫誘導以及植物與微生物互作中的功能有待于進一步深入研究。
PtPR-10內部含有一個高度保守的“P-LOOP”基序(G-x-GG-x-G-xx-K),廣泛存在于磷酸化激酶和核酸結合蛋白的結構域[39],該區域的磷酸化可能與其核酸酶活性相關[40-41]。目前,從樺樹、棉花、辣椒、刺茄和花生中分離純化的PR-10均證實具有體外核酸酶活性[42-46],而從西芹、苜蓿和馬鈴薯中分離得到的 PR-10 卻無體外核酸酶活性[47-49]。Chadha和 Das[43]研究認為,AhPR-10 蛋白通過主動運輸進入細胞后而發揮核酸酶活性降解核酸,進而殺死病原菌;此外,PR-10所包含的Bet_v_1結構域還能夠與細胞分裂素、脂肪酸和黃酮類物質相結合,進而參與類固醇激素介導的植物防御反應或生長發育調控[50-51]。本研究結果發現,PtPR-10 表達量于SA處理6h后即達到最高值,而隨后呈緩慢遞減趨勢;但PtPR-10表達水平卻隨著MeJA誘導時間的延長而不斷升高,因此,結合本實驗中PtPR-1誘導表達結果,推測PtPR-10可能只響應SA信號途徑,而不受MeJA的誘導。另外,蛋白序列比對和系統發育樹分析顯示,PR10家族的保守性不高,除了核心區域和磷酸化位點外,其余的氨基酸殘基變化差異很大,這可能是植物對多樣化的生物非生物脅迫的一種適應。
綜上所述,PtPR-1可能響應SA和JA雙重誘導,而PtPR-10可能只受SA誘導表達,PtPR-5受誘導表達并不顯著,說明病程相關蛋白基因具有不同的誘導表達模式,體現了植物體內多種信號通路調控基因表達的復雜性;而作為植物免疫系統效應因子的病程相關蛋白與抗病信號通路密不可分,是探討植物抗病機理的重要環節。因此,開展毛白楊病程相關蛋白基因的克隆,并結合信號傳導網絡對基因的功能及調控機理進行深入研究,可為林木抗病基因工程研究奠定重要基礎。
[1]Antoniw J F,White R F.The effects of aspirin and polyacrylic acid on soluble leaf proteins and resistance to virus infection in five cultivars of tobacco[J].Journal of Phytopathology,1980,98(3):331-341.
[2]Van Loon L C.The nomenclature of pathogenesis-related proteins[J].Physiological and Molecular Plant Pathology,1990,37(5):229-230.
[3]Van Loon L C,Pierpoint W S,Boller T,et al.Recommendations for naming plant pathogenesis-related proteins[J].Plant Molecular Biology Reporter,1994,12(3):245-264.
[4]Van Loon L C,Rep M,Pieterse C M J.Significance of inducible defense-related proteins in infected plant[J].Annual Review of Phytopathology,2006,44(1):135-162.
[5]Van Loon L C,Van Strien E A.The families of pathogenesis-related proteins,their activities,and comparative analysis of PR-1 type proteins[J].Physiological and Molecular Plant Pathology,1999,55(2):85-97.
[6]Jansson S,Douglas C J.Populus:a model system for plant biology[J].Annual Review of Plant Biology,2007,58(3):435-458.
[7]Zhu Z T,Zhang Z Y.The status and advances of genetic improvement of Populus tomentosa Carr[J].Journal of Beijing Forest University(English Edition),1997,6(1):1-7.
[8]Arnold K,Bordoli L,Kopp J,et al.The SWISS-MODEL Workspace:A web-based environment for protein structure homology modeling[J].Bioinformatics,2006,22(2):195-201.
[9]Guex N,Peitscn M C.SWISS-MODEL and the Swiss-PdbViewer:An environment for comparative protein modeling[J].Electrophoresis,1997,18(2):2714-2723.
[10]Schwede T,Kopp J,Guex N,et al.SWISS-MODEL:an automated protein homology-modeling server[J].Nucleic Acids Research,2003,31:3381-3385.
[11]Regier N,Streb S,Cocozza C,et al.Drought tolerance of two black poplar(Populus nigra L.)clones:contribution of carbohydrates and oxidative stress defence[J].Plant Cell Environ,2009,32(12):1724-1736.
[12]Rinaldi C,Kohler A,Frey P,et al.Transcript profiling of poplar leaves upon infection with compatible and incompatible strains of the foliar rust Melampsora larici-populina[J].Plant Physiology Preview,2007,144(1):347-366.
[13]Gattiker A,Gasteiger E,Bairoch A.ScanProsite:a reference implementation of a PROSITE scanning tool[J].Applied Bioinformatics,2002,1(2):107-108.
[14]Thompson C E,Fernandes C L,De Souza O N,et al.Molecular modeling of pathogenesis-related proteins of family 5[J].Cell Biochemistry and Biophysics,2006,44(3):385-394.
[15]Hammerschmidt R.Induced disease resistance:how do induced plants stop pathogens[J].Physiological and Molecular Plant Pathology,1999,55:77-84.
[16]Delaney T P,Friedrich L,Ryals J A.Arabidopsis signal transduction mutant defective in chemically and biologically induced disease resistance[J].Proc Natl Acad Sci U S A,1995,92(4):6602-6606.
[17]Gaffney T,Friedrich L.Requirement of salicylic acid for the induction of systemic acquired resistance[J].Science,1993,261(6):754-756.
[18]Shulaev V,Leon J.Is salicylic acid a translocated signal of systemic acquired resistance in tobacco? [J].Plant Cell,1995,7(3):1691-1701.
[19]Annemart Koornneef and Corné M J Pieterse.Cross Talk in Defense Signaling[J].Plant Physiology,2008,146(3):839-844.
[20]Dixon D C,Cutt J R,Klessig D F.Differential targeting of the tobacco PR-1 pathogenesis-related proteins to the extracellular space and vacuoles of crystal idioblasts[J].EMBO Journal,1991,10:1317-1324.
[21]Eyal Y,Meller Y,Lev-Yadun S,et al.A basic-type PR-1 promoter directs ethylene responsiveness,vascular and abscission zone-specific expression[J].The Plant Journal,1993,4:225-234.
[22]Niki T,Mitsuhara I,Seo S,et al.Antagonistic effect of salicylic acid and jasmonic acid on the expression of pathogenesis-related(PR)protein genes in wounded mature tobacco leaves[J].Plant and Cell Physiology,1998,39(2):500-507.
[23]Riviere M P,Marais A,Ponchet M,et al.Silencing of acidic pathogenesis-related PR-1 genes increases extracellular β-(1→3)-glucanase activity at the onset of tobacco defence reactions[J].Journal of Experimental Botany,2008,59(6):1225-1239.
[24]Sessa G,Yang X Q,Raz V,et al.Dark induction and subcellular localization of the pathogenesis-related PRB-1b protein[J].Plant Molecular Biology,1995,28(3):537-547.
[25]Ward E R,Uknes S J,Williams S C,et al.Goordinate gene activity in response to agents that induce systemic acquired resistance[J].Plant Cell,1991,3(3):1085-1094.
[26]Delaney T P,Uknes S,Vemooij B,et al.A central role of salicylic acid in plant disease resistance[J].Science,1994,266(2):1247-1250.
[27]Dong X N.NPR1,all things considered[J].Current Opinion in Plant Biology,2004,7(6):547-552.
[28]Mou Z L,Fan W H,Dong X N.Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes[J].Cell,2003,113(1):935-944.
[29]Spoel S H,Koornneef A,Claessens S M C,et al.NPR1 modulates cross-talk between salicylate-and jasmonate-dependent defense pathways through a novel function in the cytosol[J].Plant Cell,2003,15(2):760-770.
[30]Zhang Y L,Fan W H,Kinkema M,et al.Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene[J].Proc Natl Acad Sci U S A,1999,96:6523-6528.
[31]Petersen M,Brodersen P,Naested H,et al.Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance[J].Cell,2000,103(7):1111-1120.
[32]Creelman R A,Mullet J E.Biosynthesis and action of jasmonates in plants[J].Annual Review of Plant Biology,1997,48:355-381.
[33]Repka V,Fischerova I,Silharova K.Methyl Jasmonate is a potent elicitor of multiple defense responses in grapevine leaves and Cell-Suspension cultures[J].Biologia Plantarum,2004,48(2):273-283.
[34]Rao M V,Lee H,Creelman R A,et al.Jasmonic acid signaling modulates ozone-induced hypersensitive cell death[J].Plant Cell,2000,12(9):1633-1646.
[35]Leon Reyes A,Spoel S H,De Lange E S,et al.Ethylene modulates the role of nonexpressor of pathogenesis-related genes1 in cross talk between salicylate and jasmonate signaling[J].Plant Physiology,2009,149:1797-1809.
[36]Leon-Reyes A,Van der Does D,De Lange E S,et al.Salicylate-mediated suppression of jasmonate-responsive gene expression in Arabidopsis is targeted downstream of the jasmonate biosynthesis pathway[J].Planta,2010,232(6):1423-1432.
[37]Mao P,Duan M R,Wei C H,et al.WRKY62 transcription factor acts downstream of cytosolic NPR1 and negatively regulates jasmonate-responsive gene expression[J].Plant and cell physiology,2007,48:833-842.
[38]Midoro-Horiuti T,Brooks E C,Goldblum R M.Pathogenesis-related proteins of plants as allergens[J].Annals of Allergy,Asthma & Immunology,2001,87(4):261-271.
[39]Saraste M,Sibbald P R,Wittinghofer A.The P-loop-a common motif in ATP-and GTP-binding proteins[J].Trends in Biochemical Science,1990,15(11):430-434.
[40]Hoffmann-Sommergruber K,Vanek-Krebitz M,Radauer C,et al.Genomic characterization of members of the Bet v 1 family:genes coding for allergens and pathogenesis-related proteins share intron positions[J].Gene,1997,197(1/2):91-100.
[41]Bantignies B,Séguin J,Muzac I,et al.Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots[J].Plant molecular Biology,2000,42(6):871-881.
[42]Bufe A,Spangfort M D,Kahlert H,et al.The major birch pollen allergen,Bet v1,shows ribonuclease activity[J].Planta,1996,199(3):413-415.
[43]Chadha P,Das R H.A pathogenesis related protein,AhPR10 from peanut:an insight of its mode of antifungal activity[J].Planta,2006,225(1):213-222.
[44]Liu J J,Ekramoddoullah A K M.The family 10 of plant pathogenesis-related proteins:Their structure,regulation,and function in response to biotic and abiotic stresses[J].Physiological and Molecular Plant Pathology,2006,68:3-13.
[45]Park C J,Kim K J,Shin R,et al.Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway[J].Plant Journal,2004,37(2):186-198.
[46]Zhou X J,Lu S,Xu Y H,et al.A cotton cDNA(GaPR-10)encoding a pathogenesis-related 10 protein with in vitro ribonucle-ase activity[J].Plant Science,2002,162(4):629-636.
[47]Breda C,Sallaud C,Turk J,et al.Defense reaction in Medicago sativa:a gene encoding a class 10 PR protein is expressed in vascular bundles[J].Molecular plant-microbe interactions,1996,9(8):713-719.
[48]Liu X J,Huang B B,Lin J,et al.A novel pathogenesis-related protein(SsPR10)from Solanum surattense with ribonucleolytic and antimicrobial activity is stress-and pathogen-inducible[J].Journal of Plant Physiology,2006,163(5):546-556.
[49]Matton D P,Brisson N.Cloning,expression,and sequence conservation of pathogenesis-related gene transcripts of potato[J].Molecular plant-microbe interactions,1989,6(2):325-331.
[50]Koistinen K M,Soininen P,Venalainen T,et al.Birch PR-10c interacts with several biologically important ligands[J].Phytochemistry,2005,66(21):2524-2533.
[51]Mogensen J E,Wimmer R,Larsen J N,et al.The major birch allergen,Bet v1,shows affinity for a broad spectrum of physiological ligands[J].Journal of Biological Chemistry,2002,277(26):23684-23692.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
