北方6種綠化灌木水分脅迫的生理響應1)
2012-09-18 01:27:50柴春榮
東北林業大學學報 2012年6期
關鍵詞:質量
柴春榮
(黑龍江省科學院自然與生態研究所,哈爾濱,150040)
穆立薔
(東北林業大學)
梁 鳴
(黑龍江省科學院自然與生態研究所)
王睿姝
(東北林業大學)
我國城市干旱環境普遍存在。在這些地區發展節水型園林的主要措施之一就是大量應用耐旱植物,尤其是具有較強適應性和抗逆性的植物。城市綠化時需要優先考慮樹種的抗旱性[1]。本研究選擇6種灌木作為試驗材料,研究其在水分脅迫下的生理指標變化,對其抗旱適應性有了初步了解。結果可為城市園林樹種選擇及其應用提供參考。
1 材料與方法
紅瑞木(Cornus alba)、紅王子錦帶(Weigela florida‘Red Prince’)、麥李(Cerasus glandulosa)、遼東水蠟(Ligustrum obtusifolium)、彩葉杞柳(Salix integra‘Hakuro Nishiki’)和金銀忍冬(Lonicera maackii)2年生實生苗為試驗材料。
試驗采用盆栽控水法,每盆1株放溫室大棚內,采用稱質量法設置4種不同的土壤含水量,A處理做為對照正常澆水,B處理為輕度脅迫,C處理為中度脅迫,D處理為重度脅迫。土壤含水量分別為A處理為(50% ~55%)、B處理為(40% ~45%)、C處理為(30% ~35%)和D處理為(20% ~25%),控水處理40 d后開始采樣,對生理指標進行測定。
土壤含水量采用烘干法測定,可溶性糖質量分數的測定采用蒽酮比色法[2],丙二醛摩爾質量濃度的測定采用雙組分光光度法[3],相對電導率的測定采用電導率儀法[4],游離態脯氨酸質量分數的測定采用酸性茚三酮比色法[4],葉綠素的DSPA(Specialty Products Agricultural Division)值(即葉綠素儀通過測量葉子對兩個波長段里的吸收率,來評估當前葉子中的葉綠素的相對含量)采用DSPA-502型葉綠素儀測定。
2 結果與分析
2.1 相對電導率的變化
細胞質膜是細胞與環境之間的界面和屏障。在水分脅迫下,細胞原生質膜的結構和功能首先受到傷害,引起細胞內電解質外滲,導致組織浸出液的相對電導率增大。因此,可以通過測定組織浸出液相對電導率在水分脅迫下的高低變化來表明植物組織受傷害的程度。水分脅迫對植物的傷害,在細胞水平上以質膜相對透性的增大為主要特征,傷害越重,透性越大,外滲物越多,相對電導率的增加也愈大,抗性愈弱。質膜相對透性變化愈小,對水分虧缺的抵抗性愈好,抗旱性也愈好。綜合水分因素,可以進一步證實膜系統受損程度與葉片保水能力和抗旱性呈顯著負相關。
從表1可以看出,遼東水臘和麥李的相對電導率隨土壤含水量的降低呈先下降后上升的趨勢。方差分析顯示,各處理與A處理相比達到顯著水平,遼東水臘上升的幅度為48.39%,麥李為31.91%。其它4樹種相對電導率并沒有很大的變化,試驗期間葉片相對電導率差異不顯著,說明其它4樹種即使在長時間干旱脅迫下,細胞膜損傷也不會太嚴重。這表明其它4樹種葉片細胞膜對于干旱環境具有很強的適應性。
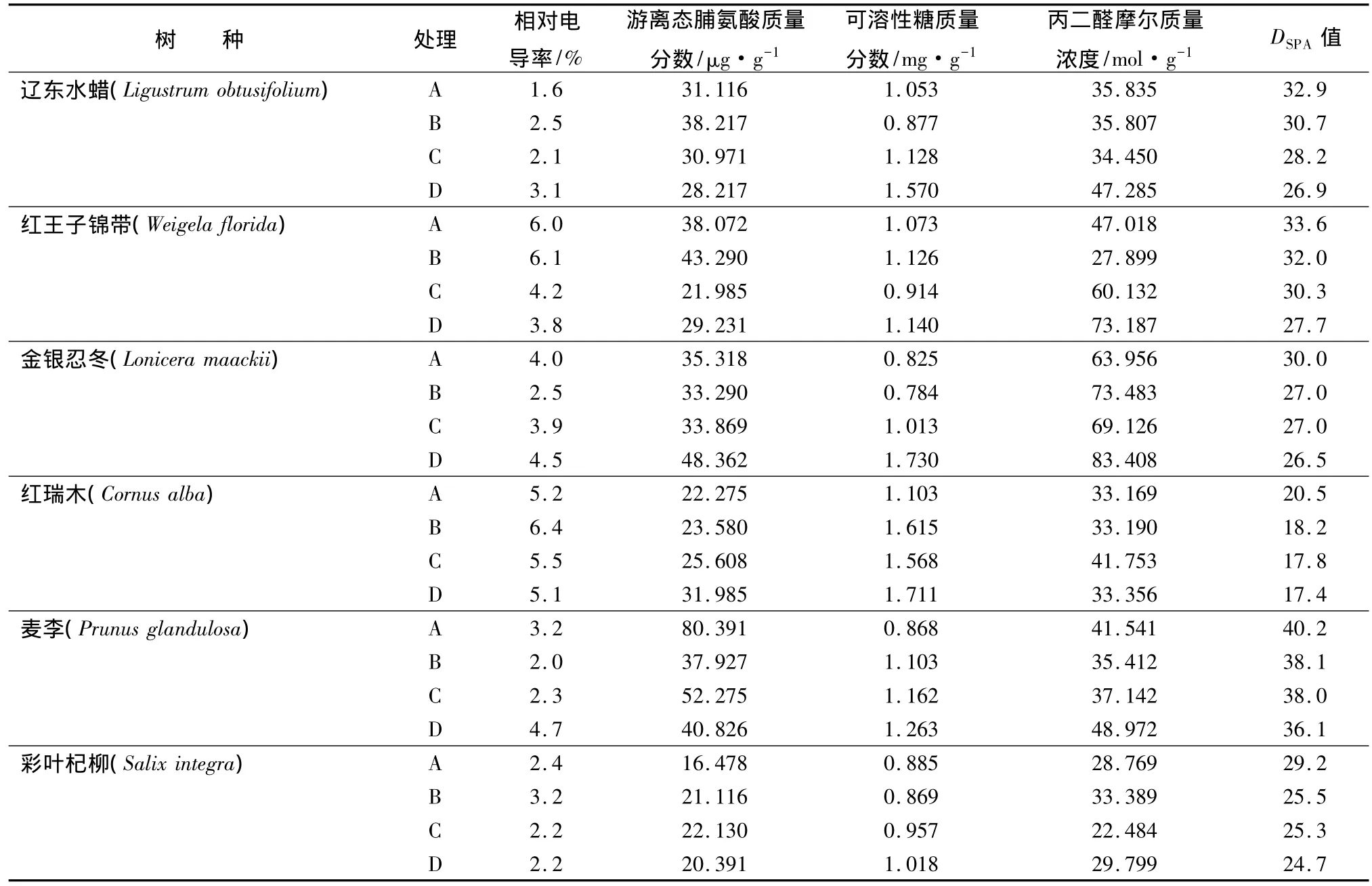
表1 水分脅迫下生理指標的變化
2.2 游離脯氨酸質量分數的變化
游離態脯氨酸是水溶性最大的氨基酸,它具有較強的水合能力,在正常情況下,植物游離脯氨酸質量分數很少,占總量的百萬分之幾,但干旱脅迫時,它可大量積累。它的增加有助于細胞或組織持水,減少脫水,維持各器官較強的滲透調節能力,從而提高植株對干旱的適應能力。
干旱脅迫使6樹種游離態脯氨酸大量生成,有的樹種游離態脯氨酸質量分數隨著干旱強度增加而一直增加;有的樹種在輕度、中度干旱下,游離態脯氨酸質量分數增加,而在重度干旱下則有降低的趨勢。
從表1可以看出,除麥李外,其它樹種在不同處理下,游離態脯氨酸質量分數都有不同程度的增加。遼東水臘和紅王子錦帶的游離態脯氨酸質量分數在B處理時上升,到了C處理、D處理又開始下降,但幅度不大。金銀忍冬、紅瑞木、彩葉杞柳的游離態脯氨酸質量分數隨土壤含水量下降呈上升趨勢,但上升幅度較小,而且水平較低。
麥李的游離態脯氨酸質量分數隨土壤含水量下降呈下降趨勢,而且各處理與A處理的差異顯著。大量研究認為植物游離態脯氨酸累積能力與品種抗旱性呈正相關[5]。游離態脯氨酸和可溶性糖是植物體內兩種重要的滲透調節物質,本試驗中遼東水臘、紅王子錦帶和麥李的游離態脯氨酸質量分數在土壤含水量降低到D處理時,都呈下降趨勢,表明遼東水臘、紅王子錦帶和麥李滲透調節能力大部分是由可溶性糖來完成的。由此可見,游離態脯氨酸累積與植物抗旱性的關系可能并不十分簡單。游離態脯氨酸的積累對植物抗旱有益,但游離態脯氨酸生物合成來源于谷氨酸;隨著干旱脅迫的發展,碳水化合物的供應受阻,將影響到谷氨酸合成,進而影響游離態脯氨酸合成。因此,有些植物游離態脯氨酸的積累最終在干旱處理末期下降和停止。
隨著土壤含水量降低,6樹種的游離態脯氨酸質量分數由高到低依次為紅瑞木、金銀忍冬、彩葉杞柳、遼東水臘、紅王子錦帶、麥李。
2.3 可溶性糖質量分數的變化
可溶性糖也是植物體內一種重要的滲透調節物質,在水分脅迫下,能增加細胞液的濃度,提高植物對水分的吸收能力及保水能力,從而有利于植物適應干旱缺水的環境。所以植物在受到水分脅迫時,可溶性糖質量分數的變化在一定程度上能反映出植物對干旱環境的適應能力[6]。干旱條件下可溶性糖對植物的保護作用已被眾多研究者證實,細胞內可溶性糖質量分數增加,能降低原生質的滲透勢,有利于細胞從外界水勢降低的介質中繼續吸水,以維持其正常的代謝活動,從而增強植物體的抗旱力。
從表1可以看出,隨著土壤含水量的降低,遼東水臘、金銀忍冬和彩葉杞柳在A處理可溶性糖質量分數開始緩慢下降,到D處理上升。方差分析表明,各處理與A處理相比達到顯著水平。這可能是因為在脅迫初期,干旱對蛋白質的抑制和降解作用大,使蛋白質含量下降;隨著脅迫的加劇,干旱對蛋白質合成的促進作用增加,進而導致了蛋白質含量上升。這說明在不同水分脅迫下,苗木體內的可溶性蛋白含量變化是不穩定的。在土壤含水量B處理條件下,紅王子錦帶、紅瑞木和麥李的可溶性糖質量分數比A處理的有所增加,增幅最大的是紅瑞木,說明,紅瑞木在B處理條件下原生質的滲透調節能力較其它5個樹種強。隨著土壤含水量的降低,6樹種的可溶性糖質量分數都呈現上升趨勢,上升幅度最大是金銀忍冬52.31%,其次為紅瑞木35.53%,遼東水臘32.93%,麥李31.27%,彩葉杞柳13.06%,上升幅度最小為紅王子錦帶5.88%。
2.4 丙二醛摩爾質量濃度的變化
植物在逆境脅迫過程中,細胞內產生過量的自由基會引發或加劇膜脂過氧化作用而造成細胞膜系統的損傷,嚴重時會導致細胞死亡。而膜脂過氧化物的最終分解產物即是丙二醛。它是膜脂過氧化的主要產物之一,是細胞中有毒性的物質,能夠引起細胞膜功能紊亂。丙二醛摩爾質量濃度的變化反映著干旱脅迫下細胞內氧自由基積累導致的脂膜過氧化程度,而脂膜過氧化程度的改變可反映植物細胞受損傷的程度[7]。
一般認為,在干旱脅迫中,植物丙二醛的摩爾質量濃度與抗旱性呈負相關,抗旱性弱的品種丙二醛摩爾質量濃度增加迅速,抗旱性強的品種則積累較少。從表1可以看出,丙二醛摩爾質量濃度隨著土壤含水量的降低都有不同程度的上升,說明了6樹種的膜系統受傷害的程度較低,受害不嚴重。遼東水臘、紅瑞木、麥李、彩葉杞柳各處理之間的丙二醛摩爾質量濃度差異不顯著,紅王子錦帶、金銀忍冬各處理之間丙二醛摩爾質量濃度差異顯著。就總體趨勢而言,隨著干旱脅迫的發生和加劇,6樹種均產生了膜脂過氧化作用,丙二醛的摩爾質量濃度均有所增大,但是各樹種的積累速度和方式不一致。遼東水臘、紅王子錦帶、麥李的丙二醛摩爾質量濃度呈先下降后上升的趨勢,特別是紅王子錦帶土,當壤含水量降低到D處理后,丙二醛摩爾質量濃度呈急劇上升的趨勢,金銀忍冬、紅瑞木、彩葉杞柳的丙二醛摩爾質量濃度均呈先上升再下降再上升的趨勢。增幅最大的是紅王子錦帶為35.76%,其次遼東水臘為24.21%,金銀忍冬為 23.32%,麥李為 15.17%,彩葉杞柳為3.46%,紅瑞木最小為0.6%。因此,6樹種的丙二醛摩爾質量濃度由低到高依次為紅瑞木、彩葉杞柳、麥李、金銀忍冬、遼東水臘、紅王子錦帶。
2.5 葉綠率DSPA值的變化
葉綠素作為光合色素中的重要色素分子,參與光合作用中光能的吸收、傳遞和轉換等過程,在光合作用中占有重要地位。一般認為,水分脅迫可導致植物葉片中葉綠素含量的降低,葉子缺水不僅影響葉綠素的生物合成,而且促進已形成的葉綠素加速分解,造成葉子發黃。SPAD-502葉綠素儀是一種輕便的葉綠素檢測儀,帶到田間可直接測定出DSPA值,DSPA值可以較好地反映植物葉片上葉綠素的含量[8]。一定范圍內葉綠素含量的高低直接影響葉片的光合能力[9]。
一般認為,葉綠素總量的變化幅度可以用作衡量植物水分脅迫敏感性強弱的重要指標,其變化較大的抗旱性相應較低。在干旱脅迫下,高的葉綠素含量能夠維持光合作用的進行,有利于增強材料的抗旱性。從表1可以看出隨土壤含水量的降低,6樹種的葉綠素DSPA值都有所下降。土壤含水量降到B處理時,6樹種的葉綠素DSPA值平均降幅為8%;當土壤含水量降到D處理時,6樹種的葉綠DSPA值平均降幅為15%。遼東水蠟和紅王子錦帶的葉綠素DSPA值降幅最大,均下降18%;其次為紅瑞木和彩葉杞柳,均下降15%,金銀忍冬下降12%,麥李的降幅最小為10%。當干旱脅迫時間延長或加重,葉綠素的含量呈現下降趨勢。這可能是因為干旱脅迫的加劇引起植物細胞內生理、生化改變,使葉綠素合成受阻,降解加快,葉綠素含量迅速下降[10]。
2.6 水分脅迫下6樹種生理特性的綜合評價
樹木耐旱能力的強弱不是一個絕對概念,其排序只是相對而言,由于實驗的材料和方法不同所得出的結論也會出現一些差異。需要說明的是苗木的耐旱性受到多個抗旱指標的影響,用不同的抗旱指標對其進行評價得出的結論不同。上述各生理指標的變化趨勢,僅是在試驗范圍內的結論,當水分脅迫超過一定限度時,其變化趨勢可能發生改變[11]。本研究選擇相對電導率、游離態脯胺酸質量分數、可溶性糖質量分數、丙二醛摩爾質量濃度、葉綠素DSPA值等指標,采用隸屬函數法對6樹種抗旱性進行綜合評價。
評定方法采用模糊數學反隸屬函數方法來計算[12]。計算公式為:
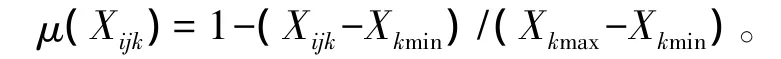
式中:μ(Xijk)為第i樹種在第j個土壤水分處理對于第k項水分參數的隸屬函數值;Xijk為第i個樹種第j個土壤水分處理第k個水分參數測定值;Xkmax、Xkmin為6樹種第k項指標的最大值和最小值;μ(Xijk)值越大,耐旱性越強。先分別計算5項水分參數隸屬函數值。
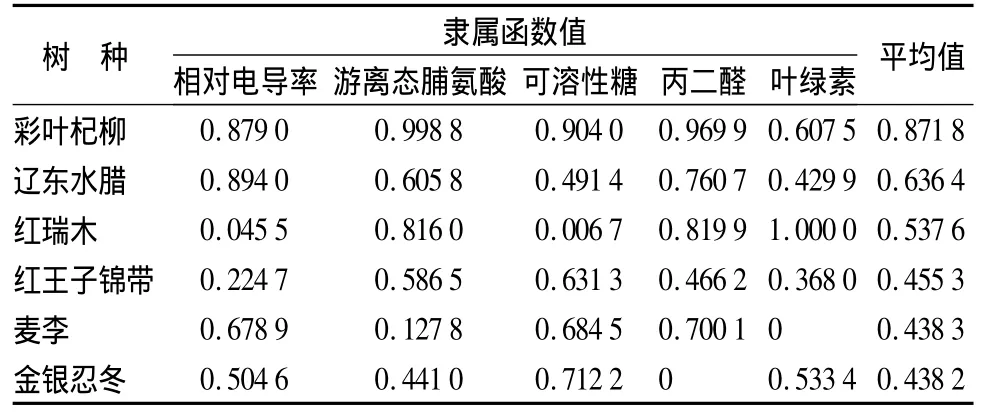
表2 6樹種抗旱能力的綜合評價
表2為6樹種抗旱能力的綜合評判結果,可以看出6樹種的抗旱性有強弱差別,抗旱能力大小依次為:彩葉杞柳>遼東水蠟>紅瑞木>紅王子錦帶>麥李>金銀忍冬。
3 結論與討論
本試驗中,6樹種的游離態脯氨酸質量分數有的先下降再上升,有的則呈現下降上升再下降的變化。隨土壤含水量下降,金銀忍冬、紅瑞木、彩葉杞柳的降游離態脯氨酸質量分數呈上升趨勢,但上升幅度緩慢,且水平較低;遼東水臘、紅王子錦帶、麥李的游離態脯氨酸質量分數都呈下降趨勢。由此可見,游離態脯氨酸累積與植物抗旱性有密切的關系,但并不簡單還有待進一步研究。
植物在受到水分脅迫后,抵御適應干旱的途徑和方式多種多樣,因而用單一或個別生理生化指標難以進行復雜的抗旱鑒定,所以應采用多項指標進行綜合評價。植株葉片相對電導率、游離脯氨酸、可溶性糖、丙二醛和葉綠素含量均是植物生理生化信息的反映,在干旱脅迫下,遼東水臘和麥李的相對電導率呈上升趨勢,上升幅度分別為48.39%和31.91%,其它4樹種相對電導率上升慢,且變化不明顯,組織或細胞受損輕;游離脯氨酸的積累能增加細胞滲透勢和親水,有利于組織或細胞的保水;細胞內可溶性糖質量分數增加,能降低原生質的滲透勢,有利于細胞從外界水勢降低的介質中繼續吸水,以維持其正常的代謝活動,從而增強植物體的抗旱力;丙二醛摩爾質量濃度隨著土壤含水量的降低都有不同程度的上升,說明了6樹種的膜系統受傷害的程度較低,受害不嚴重;6樹種的葉綠素DSPA值都有所下降,這可能是因為干旱脅迫的加劇引起植物細胞內生理、生化改變,使葉綠素合成受阻,降解加快,葉綠素含量迅速下降。綜合多項指標比較結果,6樹種的抗旱能力大小依次為:彩葉杞柳>遼東水蠟>紅瑞木>紅王子錦帶>麥李>金銀忍冬。
[1]陳娟,陳其兵,潘遠志,等.6種野生灌木的抗旱性的研究[J].四川林業科技,2007,28(5):50-54.
[2]王晶英,敖紅,張杰,等.植物生理生化實驗技術與原理[M].哈爾濱:東北林業大學出版社,2003:11-13.
[3]高俊鳳.植物生理學實驗指導[M].北京:高等教育出版社,2006:210-211.
[4]鄒琦.植物生理生化實驗指導[M].北京:中國農業出版社,1995.
[5]周源.七種鳶尾的抗旱性研究[D].烏魯木齊:新疆農業大學,2008.
[6]邵麟惠.柴達木盆地六種灌木抗旱性綜合評價研究[D].蘭州:甘肅農業大學,2007.
[7]趙明范,葛成,翟志中.干旱地區次生鹽堿地主要造林樹種抗鹽指標的確定及耐鹽能力排序[J].林業科學研究,1997,10(2):194-198.
[8]孫海鋒.干旱脅迫對新疆不同基因型春大豆光合生理響應的抗旱研究[D].石河子:石子河大學,2008.
[9]張永強,毛學森,孫宏勇,等.干旱脅迫對冬小麥葉綠素熒光的影響[J].中國生態農業學報,2002,10(4):13-15.
[10]趙瑾,白金,潘青華,等.干旱脅迫下圓柏不同品種(系)葉綠素含量變化規律[J].中國農學通報,2007,23(3):236-239.
[11]孫志虎,王慶成.應用PV技術對北方4種闊葉樹抗旱性的研究[J].林業科學,2003,29(2):33-38.
[12]陳榮敏,楊學舉,梁鳳山,等.利用隸屬函數法綜合評價冬小麥的抗旱性[J].河北農業大學學報,2002,25(2):7-9.
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
