尾巨桉人工林營養元素積累及其生物循環特征1)
2012-09-18 01:27:50朱宇林
東北林業大學學報 2012年6期
關鍵詞:質量
朱宇林
(玉林師范學院,玉林,537000)
何 斌 楊鈣仁 羅柳娟 劉紅英 劉 莉 覃祚玉
(廣西大學)
桉樹(Eucalyptus)具有適應能力強、生長迅速、單產高、生長周期短、樹干通直、用途廣泛和經濟效益高等特點,為世界許多國家引種和推廣的速生樹種[1-2],已成為我國華南地區短周期工業用材林生產基地的首選樹種。尾巨桉(Eucalyptus urophylla×E.grandis)是由尾葉桉(E.urophylla)和巨桉(E.grandis)雜交獲得的雜交種,具有生長快、干形好、輪伐期短、伐根萌芽力強、無性繁殖容易、經濟效益高等特點,是目前我國南方實施速生豐產用材林基地建設的主要桉樹優良品種之一,并取得了顯著的經濟效益[3-4]。但是,如何合理經營和科學地管理尾巨桉人工林,維持其持久生產力,已成為當前和今后林業生產實踐和生態環境建設面臨的實際問題。
養分循環是維持森林生態系統穩定和生產力的功能過程之一[5-6],尾巨桉林養分循環的研究不僅能揭示該生態系統物質循環機制,而且對指導林業生產、調節和改善林木生長的環境、提高系統的養分利用率和最大限度地提高生產力都具有重要意義。國內有關桉樹人工林養分循環的研究已有不少報道[7-12],但多數都是對某一年齡(或林齡)桉樹養分循環進行研究,其中對尾巨桉人工林的研究僅見于幼齡林(2年生)的報道[12],缺乏對林分養分循環的連續監測與分析,難以反映尾巨桉人工林生長中的養分循環過程。為此,本研究通過4.5 a的定位觀測與分析,對1.5~4.5年生尾巨桉人工林主要營養元素的積累、分布和生物循環進行了研究,以揭示尾巨桉人工林營養元素循環的特點及其變化趨勢,為桉樹生產的經營管理,特別是林木施肥管理,以及防止桉樹人工林地力衰退提供科學依據。
1 試驗地概況
試驗地位于廣西南寧市北郊,地理位置為東經108°21',北緯 22°58',地處南寧盆地的北緣,大明山山脈南伸的西支。屬南亞熱帶季風氣候,年平均氣溫21.8℃,極端最高氣溫40℃,極端最低氣溫-2℃,≥10℃年積溫約7200℃。年降水量1200~1500 mm,降雨多集中在5—9月份,年蒸發量1250~1620 mm,年日照時數1450~1650 h,相對濕度大約79%[5]。標準地位于山坡中部,海拔高度約200 m,坡度25°~28°,東南坡,土壤類型為砂頁巖發育形成的赤紅壤,土壤厚度在80 cm以上。
試驗地前茬林分均為杉木(Cunninghamia lanceolata)純林,于2001年年底砍伐,經煉山后按株行距2 m×3 m挖明穴,穴的長×寬×高為60 cm×40 cm×40 cm。于2002年4月用尾巨桉營養苗栽植,初植密度為1650株/hm2,種植前0~40 cm土層土壤有機質、全N、全P和全K質量分數分別為16.72、0.97、0.32 和14.20 g/kg,交換性鈣和交換性鎂質量分數分別為49.52和7.86 μg/g。造林前每穴施鈣鎂磷肥500.0 g作基肥,植后當年7月份鋤草撫育并施尿素100.0 g,9月份鋤草撫育并施復合肥250.0 g。經林分自疏和間伐后,各不同林齡尾巨桉人工林林分特征見表1(均為當年10月份測定的數據)。

表1 尾巨桉人工林林分特征
2 研究方法
標準地的設置和林分生物量的測定:在立地條件相似的尾巨桉人工林中設置3塊20 m×20 m固定標準地,并對標準地內的每株樹木進行編號,每年10月測定標準地內樹木的樹高和胸徑。根據林分生長調查結果,同時為了不破壞試驗地的林分,每年均在標準地外圍選擇代表平均值的3株標準木(平均木),采用收獲法測定標準木的生物量。地上部分采用Monsic分層切割法,每2 m為一區分段,分干材、干皮、樹枝、樹葉,分別稱質量;地下部分(根系)采用全根挖掘法,分根蔸、粗根(根系直徑(d)≥2.0 cm)、中根(0.5≤d<2.0 cm)、細根(d<0.5 cm),分別稱質量。取樣測定各器官的含水率及干質量,計算林分生物量,以年平均增長量作為凈生產力的估測指標。同時在每個標準地內各隨機設置5個1 m×1 m的小樣方,調查樣方內植物種類、個體數、高度和覆蓋度等。灌木和草本層用樣方法收割和稱量地上部鮮質量,再挖掘和稱量樣方地下20.0 cm范圍內的根;凋落物層則采用收集和稱量樣方內所有凋落物,同時取樣測定含水率和干質量。
年凋落物歸還量的測定:在每個標準地內隨機設置5個1 m×1 m的木框架尼龍網收集器,收集網的孔徑為1.0 mm,每月月底收集凋落物1次,帶回室內,按葉、枝、花果和雜物等組分烘干測定生物量[6]。
植物樣品營養元素分析:在測定生物量的同時按不同組分采集分析樣品;凋落物樣品為按各月份凋落物質量比例各選取一定量的凋落物混合后作為化學分析樣品。樣品中N質量分數采用濃H2SO4-HClO4-氨氣敏電極法測定[13],P質量分數用鉬銻抗比色法測定,K用火焰光度計法測定,Ca、Mg用原子吸收光譜法測定[14]。
養分循環參數的計算:采用養分利用系數、循環系數和周轉時間等生物循環參數來分析養分循環的特征。按照生物循環公式(吸收量=存留量+歸還量)進行計算。上述養分利用系數為吸收量與儲存量的比值,表明林木維持其生長所需的營養元素量;循環系數為歸還量與吸收量的比值,表征營養元素的循環強度;周轉時間為營養元素經歷一個循環周期所需的時間,即營養元素的總儲存量除以歸還量[15]。
3 結果與分析
3.1 營養元素質量分數
由表2可見,尾巨桉平均單株不同器官的營養元素質量分數相差較大,并且在不同林齡之間也存在一定的差異。作為同化器官的樹葉,其生長周期短,是合成有機物質的場所,也是代謝最活躍的器官,因此,其營養元素質量分數大多為最高;而干材以木質為主,其生理功能最弱,大多數養分已被消耗或轉移,因而元素質量分數也最低。就尾巨桉各器官中營養元素質量分數而言,其排序大致為:樹葉>干皮>樹枝>樹根>干材。在不同林齡的人工林中,各器官營養元素質量分數也有所差異。除N元素外,干材和樹根,尤其是干材的不同營養元素質量分數表現出隨林齡增長而減小的趨勢。在不同器官中,N在樹葉中的質量分數最高,K、Ca和P在樹葉和干皮中的質量分數最高,Mg元素則以樹皮中的質量分數為最高。

表2 不同林齡尾巨桉人工林營養元素質量分數
3.2 營養元素積累與分布
從表3 可以看出,1.5、2.5、3.5 和 4.5 年生尾巨桉人工林營養元素積累量分別為452.19、701.52、771.89 和 850.54 kg/hm2,營養元素積累量隨林齡的增加而增大,但其增加幅度隨林齡增大而減少,其中 1.5 ~2.5 年生增加249.33 kg/hm2,增加幅度達到55.14%,遠高于其他林齡間的增長幅度。喬木層作為林分營養元素的主要組成部分,其營養元素積累量分別占不同林齡林分營養元素積累總量的 70.92%、80.16%、80.73% 和 79.74%,林下植被即草本層和灌木層分別占22.99%、14.61.%、14.69%和13.84%,林分現存地表凋落物層分別占6.12%、5.27%、4.63%和 6.48%。
如將林木器官分為樹冠(樹枝和樹葉)、樹干(干材和干皮)和樹根3部分,則尾巨桉人工林上述3個部分營養元素積累量分別占喬木層積累量的32.23% ~37.37%、49.49% ~54.44% 和 8.20% ~18.28%。而從喬木層各種營養元素積累量來看,多數以Ca積累量最大,其占總積累量的25.67% ~32.30%;P 的積累量最小,僅占3.25% ~3.99%,其它營養元素所占比例均為K>N>Mg,與位于相同區域馬占相思和厚莢相思等固氮樹種存在較大差異[6,16]。
3.3 營養元素年凈積累量
從表4 可以看出,1.5、2.5、3.5 和 4.5 年生尾巨桉人工林喬木層營養元素年凈積累量分別為213.79、224.93、178.04 和 150.72 kg/(hm2·a)。可見,種植后1.5~2.5年生是尾巨桉人工林生長最快的時期,其營養元素的凈積累量增幅也最大,隨后2.5~4.5年生營養元素的凈積累量呈下降趨勢。而從不同器官營養元素年凈積累量來看,干材營養元素年凈積累量隨林齡的增加而減少,而樹葉和干皮除1.5 ~2.5 年生呈上升外,2.5 ~3.5 年生和3.5 ~4.5年生均隨林齡增加而下降。可見,尾巨桉人工林營養元素年凈積累量的變化有利于減輕其在2.5年生后林木生長對營養元素的依賴。

表3 不同林齡尾巨桉人工林營養元素的積累與分配
不同林齡尾巨桉人工林營養元素年凈積累量多呈現出 K 或 Ca>N>Mg>P,其中1.5年生表現為 K>Ca>N>Mg,此后均為 Ca>K>N>Mg。從各營養元素在不同器官的年凈積累量看,N在樹葉和干皮中積累量最大;P在干材中積累量最大,在樹枝、樹根中積累量較小;K在干材中積累量最大,在樹枝和干皮中積累量較小;Ca和Mg在干皮中積累量最大,在干皮或樹根中積累量較小。

表4 尾巨桉人工林營養元素年凈積累量
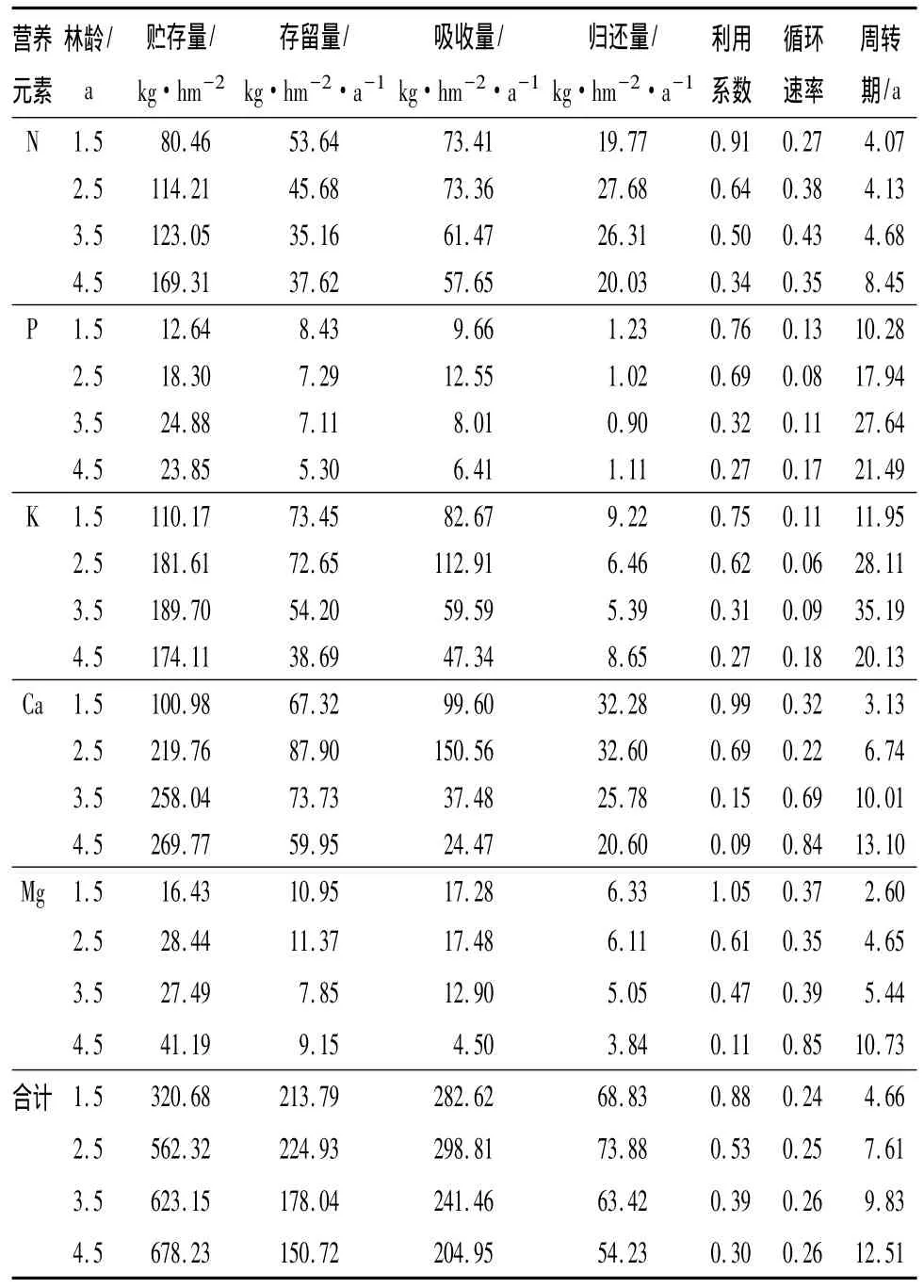
表5 尾巨桉人工林營養元素生物循環
3.4 營養元素的生物循環
從表5 可見,1.5、2.5、3.5 和 4.5 年生尾巨桉人工林5種營養元素歸還量分別為68.83、73.88、63.42和54.23 kg/(hm2·a),吸收量分別為282.61、298.81、241.461 和 204.95 t/(hm2·a),兩者均隨林齡的增長呈現先增大(1.5~2.5年生)后減少(2.5~4.5年生)的趨勢,但歸還率都不高,僅為21.46% ~26.46%。不同營養元素的年歸還量和吸收量存在較大的差異,以Ca歸還量最多,其次是N和K,P最小,且N、Ca的歸還量都與營養元素總歸還量的變化趨勢相一致,P和K的歸還量則呈現先減少(1.5~2.5年生)后增大(2.5 ~4.5 年生)的趨勢。不同林齡尾巨桉人工林存留量大致以Ca最多,其次是K、N,P最少。
不同林齡尾巨桉人工林的營養元素利用系數為0.30~0.88,隨林齡的增加而減少(表5);循環系數為0.24 ~0.26,周轉期為4.66 ~12.51 a,均隨林齡的增加而增大,其中各林齡的周轉期均明顯大于其相應林齡。不同營養元素的循環速率以Mg最快,利用系數也最高,周轉期最短;P和K的循環速率最低,利用系數相應也最低,周轉時間則最長。
4 結論與討論
尾巨桉各器官營養元素質量分數因器官和林齡不同存在差異,總體表現為樹葉>干皮>樹枝>樹根>干材。不同營養元素在各器官中的質量分數也因器官不同而存在一定的差異,N元素在樹葉中的質量分數最高,K、Ca和P元素在樹葉和樹皮中的質量分數最高,Mg元素在樹皮中的質量分數最高。
1.5~4.5年生尾巨桉人工林營養元素積累量為452.19 ~850.54 kg/hm2,其中喬木層營養元素積累量占70.92% ~80.73%,樹干(干材和干皮)營養元素積累量則占喬木層的49.49% ~54.44%。因此,從營養元素積累與分布看,在采伐時如果只取走主干,而將其他剩余物(葉、枝、根等)歸還土壤,會明顯減少林地養分的消耗,養分損失不會太大。但目前各地多數采用的是桉樹全樹利用方式,加上采伐后采用煉山整地方式,雖然提高了木材利用率,有利于營林作業,但對維持林地地力不利。
1.5、2.5、3.5 和4.5 年生尾巨桉人工林喬木層營養元素年凈積累量分別為 213.79、224.93、178.04 和150.72 kg/(hm2·a),其變化趨勢與喬木層營養元素積累量存在明顯的差異,表明種植后至2.5年生既是尾巨桉的快速生長期,同時也是其對營養元素吸收的旺盛期;由于不同林齡尾巨桉人工林營養元素年吸收量和存留量(年凈積累量)為K或Ca>N>Mg>P,而林地土壤交換性鈣、鎂含量均不高,因此,在重視N、P、K肥施用的同時,應重視Ca、Mg,特別是Ca肥的施用,以促進林木的生長發育,維持林地的養分平衡。
1.5~4.5年生尾巨桉人工林大量元素歸還量為54.23 ~73.88 kg/(hm2·a),吸收量為 204.95 ~298.81 t/(hm2·a),歸還率為 21.46% ~26.46%,遠低于相同或相近林齡的馬占相思、厚莢相思人工林(45% ~ 49%)[6,16],這也是尾巨桉人工林與杉木人工林一樣不利于土壤肥力恢復和維持的重要原因之一[17]。不同林齡尾巨桉人工林營養元素利用系數為0.30~0.88,隨林齡的增加而減少,循環系數為0.24 ~0.26,周轉期為 4.66 ~12.51 a。總的來說,1.5~4.5年生尾巨桉人工林處于生長旺盛期,營養元素的積累速率快,會大量消耗林地養分,其歸還的養分又較少,歸還速率較慢,而目前經營尾巨桉人工林的輪伐期多數為5~6 a,有的甚至縮短至不到4 a,加上多數采用桉樹全樹利用,以及采伐后采用煉山整地方式,不利于林地土壤肥力的恢復、維持和提高。因此,進行合理的林地土壤管理,尤其是進行合理施肥管理,保留林地凋落物,適當保留草本、灌木[18],或者采用桉樹與固氮樹種如相思樹混交[19-20]方式進行經營,增加地被層促進桉樹林地形成良性的物質循環,從而恢復和維持其地力,對尾巨桉人工林長期生產力的維持,促進尾巨桉人工林可持續發展有著重要的作用。
[1]殷亞方,姜笑梅,呂建雄,等.我國桉樹人工林資源和木材利用現狀[J].木材工業,2001,15(5):3-5.
[2]何斌,溫遠光,周敏毅.新栽培區尾葉桉人工林營養元素積累與分配特征[J].中南林業科技大學學報,2008,28(5):11-15.
[3]陳少雄,楊建林,周國福.不同栽培措施對尾巨桉生長的影響及經濟效益分析[J].林業科學研究,1999,12(4):357-362.
[4]朱宇林,溫遠光,曹福亮,等.短周期尾巨桉連栽林分生產力的研究[J].江西農業大學學報,2006,28(1):90-94.
[5]Sharma J C,Sharma Y.Nutrient cycling in forest ecosystems:A review[J].Agricultural Reviews,2004,25(3):157-172.
[6]何斌,余浩光,劉運華,等.不同年齡階段馬占相思(Acacia mangium)人工林營養元素的生物循環[J].生態學報,2007,27(12):5158-5167.
[7]徐大平,曾育田,李偉雄.尾葉桉幼林地上部分生物量及養分循環的研究[J].林業科學研究,1994,7(6):600-605.
[8]余雪標,白先權,徐大平,等.不同連栽代次桉樹人工林的養分循環[J].熱帶作物學報,1999,20(3):60-66.
[9]李躍林,李志輝,謝耀堅.巨尾桉人工林養分循環研究[J].生態學報,2001,21(10):1734-1740.
[10]林德喜,劉開漢,羅水發.尾葉桉營養元素動態和循環分析[J].應用與環境生物學報,2002,8(2):148-153.
[11]廖觀榮,鐘繼洪,李淑儀,等.桉樹人工林生態系統養分循環與平衡研究:Ⅱ.桉樹人工林生態系統的養分循環[J].生態環境,2003,12(3):296-299.
[12]蘇有文,何斌,張偉,等.尾巨桉人工幼林營養元素的生物循環[J].安徽農業科學,2009,37(26):12773-12775.
[13]何斌.植物氮、磷、鉀的自控遠紅外快速聯合消化和測定法[J].理化檢驗:化學分冊,1992,28(4):44-45.
[14]魯如坤.土壤農業化學分析方法[M].北京:中國農業科學技術出版社,1999:308-322.
[15]何斌,韋善華,張偉,等.黑木相思人工林營養元素生物循環特征[J].東北林業大學學報,2012,40(2):9-12,29.
[16]秦武明,何斌,覃世贏,等.厚莢相思人工林營養元素生物循環的研究[J].水土保持學報,2007,21(4):103-107.
[17]Ma Xiangqing,Heal K V,Liu Aiqin,et al.Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China[J].Ecology and Management,2007,243:61-74.
[18]Xu D,Dell B,Yang Z,et al.Effects of phosphorus application on productivity and nutrient accumulation of a Eucalyptus urophylla plantation[J].Journal of Tropical Forest Science,2005,17(3):447-461.
[19]David I,Forrester J B,Annette L C,et al.Mixed-species plantations of Eucalyptus with nitrogen-fixing trees:A review[J].Forest Ecology and Management,2006,233:211-230.
[20]姚慶端,鄒國明,王炳南,等.巨尾桉混交林生長量及土壤肥力研究[J].桉樹科技,2006,23(1):1-5.
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
