分泌性免疫球蛋白A與腸道黏膜免疫的關系及其分泌的營養調控
2012-09-11 07:35:10王定越徐盛玉楊雙源
動物營養學報 2012年10期
關鍵詞:小鼠
王定越 徐盛玉 宋 潔 楊雙源 吳 德
(四川農業大學動物營養研究所,動物抗病營養四川省重點實驗室,雅安 625014)
分泌性免疫球蛋白A(SIgA)是20世紀60年代初在人的外分泌液中發現的一種免疫球蛋白A(IgA)抗體,主要存在于唾液、乳汁、胃腸液和呼吸道分泌液等外分泌液中,為黏膜免疫的主要抗體。SIgA抗體作為黏膜表面免疫排異和抗原的有效因子,在上皮屏障中通過選擇性的黏附小腸淋巴Peyer氏結(PP)的M細胞起著免疫平衡的作用。因此,研究SIgA的功能及營養對其調控作用有著重要的現實意義。本文就SIgA、腸道黏膜免疫、SI-gA在腸道黏膜免疫中的功能及營養因子對SIgA分泌的調控作用作一綜述。
1 SIgA
腸道SIgA起始于腸黏膜固有層IgA漿細胞。IgA漿細胞在黏膜淋巴濾泡中發育,多沿上皮層分布,彌散于黏膜下層。IgA漿細胞產生11S IgA,同時胞漿內合成J鏈(含胱氨酸較多的酸性蛋白質),將漿細胞產生的IgA聚合形成多聚體IgA。基因敲除J鏈小鼠試驗發現IgA不能被多聚免疫球蛋白受體(plgR)上皮細胞有效轉運,說明J鏈參與了IgA的轉運[1-2]。多聚體IgA通過plgR的轉運作用釋放到外分泌液中,釋放出的IgA再與上皮細胞分泌的分泌片段(SC)結合后形成SIgA,最后與上皮細胞緊密連接在一起,分布在黏膜或漿膜表面發揮免疫作用[3]。
SIgA中的 IgA主要有2種類型,IgA1和IgA2,二者差異在于IgA2的鉸鏈區較IgA1缺乏13個脯氨酸富集的氨基酸殘基[3]。IgA2含有3個亞型,分別為 IgA2m(1)、IgA2m(2)、IgA2m(3),其中 IgA2m(3)是 IgA2m(1)和 IgA2m(2)的Cα區域基因移位或組合的雜交分子。IgA由3個主要區域(Cα1、Cα2和 Cα3)和各自的鉸鏈區組成。Cα3區域額外含有1個18個氨基酸的片段,被稱為尾巴片段,該片段可確保二聚體IgA和J 鏈的連接[3]。
2 腸道黏膜免疫
腸道黏膜是動物機體和內、外環境進行交流的一個大面積場所,也是病原體入侵動物的主要門戶,為了抵抗病原體的入侵,黏膜組織內的淋巴組織和免疫活性細胞共同形成的一個完整的免疫應答網絡,被稱為黏膜免疫系統[4]。根據功能和分布,黏膜免疫系統分為黏膜相關淋巴組織和彌散的免疫細胞。黏膜相關淋巴組織包括:腸道PP、孤立淋巴濾泡和腸系膜淋巴結;彌散的免疫細胞包括:腸道上皮細胞、腸道上皮內淋巴細胞、黏膜固有層漿細胞、黏膜固有層淋巴細胞。
腸道黏膜免疫系統中大部分淋巴組織和免疫細胞與SIgA存在一定的聯系,如固有層內的淋巴細胞既有T細胞又有B細胞[5],B淋巴細胞主要由小淋巴細胞和漿細胞組成,其中75% ~85%的漿細胞能夠產生SIgA。
3 SIgA在腸道黏膜免疫中的功能
3.1 維持黏膜內環境穩態
SIgA的分泌有助于宿主-寄生蟲共生、自我平衡及控制細菌在腸道內定植的程度,要確保共生菌的平衡,IgA分泌細胞需持久的慢性刺激[6]。基因敲除小鼠由于缺乏激活誘導重組IgA產生的脫氨酶,表現出腸道菌群的異常增殖[7]。可見SIgA對維持腸道菌群平衡有重要作用。
體內、外研究發現PP中樹狀突細胞(DC)可吞噬SIgA[8],SIgA通過PP進入黏膜在保護腸道屏障完整性上有著非常重要的免疫調節作用,SIgA與極化的Th2和Th3細胞一起足以調控腸道黏膜的內環境穩態。因此,SIgA也被認為是維持腸道屏障的重要調節因子[8]。缺乏plgR鼠的試驗同樣發現SIgA參與維持腸道黏膜的穩定性[9]。
由此說明,SIgA對維持腸道菌群平衡及腸道黏膜穩定性有重要作用。
3.2 干擾病菌與上皮細胞受體結合
當腸黏膜受到不同程度侵襲(病原菌、病原顆粒、細菌毒素等)時,黏膜便產生大量的SIgA,以抵抗各種病原體的入侵。SIgA發揮干擾作用的第1步是通過與病原體和毒素作用的上皮細胞受體結合,從而抗感染。SIgA通過抗體的可變區(Fv)-依賴以及Fv-非依賴機制封閉上皮細胞與病原體和毒素結合的受體。
3.2.1 Fv-依賴干擾
以霍亂毒素(CT)與SIgA的相互作用為例子。生產抗CT的二聚體IgA單克隆抗體,在體外培養的單層T84細胞中抗體均表現出通過黏附細胞頂端表面受體封阻了CT的入侵[10]。新生小鼠被動接受單克隆抗體對CT引起的分泌性腹瀉、體重降低和死亡都有免疫作用[11],由此說明,機體對CT的免疫需要SIgA,這種保護調節主要是通過抗體封閉毒素結合的上皮細胞受體而實現。
此外,SIgA還可通過阻止病毒黏附上皮細胞而防止病毒感染。通過小鼠口服接種呼腸孤1型病毒(T1L)考察單克隆IgA抗體對病毒表面抗原(黏附素和衣殼蛋白)的直接抵抗能力,結果發現,1E1(自制IgA抗體)單克隆抗體具有保護作用[12]。1E1可識別抗原決定簇的黏附素纖維蛋白——σ1蛋白并與之結合,這種蛋白能促進病毒附著于各種不同類型上皮細胞,包括M 細胞[13]。對其他病毒表面抗原,如衣殼蛋白,單克隆IgA抗體則不能通過類似與T1L相互結合的方式發揮保護作用[12]。由此表明,SIgA抗體防護腸病毒,如呼腸孤病毒最有效的方式是直接抵抗病毒表面的抗原決定簇使其不能附著于上皮細胞受體。
以上研究說明,SIgA通過Fv-依賴方式抵抗病原菌入侵時,既可以封閉毒素結合的上皮細胞受體,也可以直接抵抗病毒表面的抗原決定簇。
3.2.2 Fv-非依賴干擾
從結構上講,IgA的H鏈(FCA)和SC大部分糖基化,SIgA寡糖側鏈與管腔內的腸上皮細胞高度的相似,因而IgA和SC可以有效地作為競爭性抑制劑抑制病原體與宿主細胞的結合[14-16]。
蓖麻毒素的研究有效證實了SIgA的這種Fv-非依賴阻斷病源作用[14]。此外,SC憑借其N-和O-鏈接的碳水化合物側鏈,足以抑制毒素與受體的結合。SC可能是其他病原體,包括腸道大腸桿菌的競爭性受體[16]。SIgA的Fv-非依賴阻斷即不依賴Fv而通過結構的相似性競爭防止病原體和毒素附著上皮細胞表面。
3.3 免疫排異清除病原體
免疫排異是SIgA阻止微生物黏附、定植和侵入上皮細胞的最主要機制[17]。SIgA在腸道中識別病毒和細菌表面的多價抗原,在腸腔內凝集病原微生物進而阻止或消除它們黏附和滲透進入上皮細胞。此外,覆蓋在上皮細胞表面的黏液層不僅可以限制SIgA免疫凝集物的彌散,同時也可網羅這些凝聚物并促進它們通過腸蠕動的方式清除出腸道[18]。
3.4 降低病原菌毒力
研究發現,SIgA介導的對致命腸道病菌防護,如鼠傷寒沙門氏菌和腸炎沙門菌,既不能用封閉受體,也不能用免疫排異的機制進行解釋。鼠傷寒沙門氏菌接觸腸腔內表面后,細菌蛋白質進入宿主細胞的細胞質中,從而導致細胞骨架重排[19]。腸黏膜感染該菌后也表現出強烈的炎癥反應[20]。
鼠傷寒沙門菌中添加抗鼠傷寒沙門菌O-抗原的特異性抗體(Sal4),發現細菌運動的時間和Sal4呈劑量依賴性[21]。Sal4處理的細菌完全“癱瘓”,且Sal4的抑制作用非常快速(小于15 min)。由此表明,Sal4是細菌運動的強效抑制劑。掃描電子顯微鏡觀察對照和Sal4處理的細菌,發現對照組細菌表面均勻布滿了凹槽和凸起,而Sal4組菌膜表面平滑且有水泡樣凸起[22]。表明Sal4與O-抗原結合,擾亂細菌固有組織脂多糖層,誘導細菌菌膜表面拓撲結構的變化,從而破壞細菌外膜的完整性。
由此說明,SIgA防護O-抗原病毒感染主要是降低菌體的運動力,破壞菌膜完整性進而降低病菌的毒力。
4 營養因子對SIgA分泌的調控作用
4.1 脂肪酸
小鼠出生至斷奶(21日齡)及斷奶后1周,每天每只鼠灌喂共軛亞油酸(CLA)1.5 mg(據1%CLA飼糧換算)(Sog+EI組),第28天時屠宰,通過RT-PCR檢測發現Sog+EI組小腸和結腸IgA的相對表達量較出生至斷奶灌喂CLA(Sog組)、斷奶后1周飼糧添加1%CLA(EI組)和無CLA添加(Ref組)分別高出約6倍和4倍,進一步通過ELISA檢測腸道中SIgA的含量發現,Sog+EI組顯著高于 Sog、EI和 Ref組[23]。可見小鼠哺乳期及早期補充CLA可促進腸道SIgA的分泌,提高其腸道免疫力。
另有研究發現,小鼠空腸灌注油酸(長鏈脂肪酸)微粒可顯著提高腸腔中SIgA的含量,但當十二指腸灌服油酸微粒時卻顯著降低了SIgA的分泌。對于辛酸(中鏈脂肪酸)2種處理方式均不會引起SIgA分泌的改變[24]。
盡管已有研究發現部分脂肪酸有促進SIgA分泌的作用,然而這些脂肪酸提高SIgA分泌,促進動物腸道免疫力增強的具體途徑或機制還有待進一步研究。
4.2 氨基酸
研究發現,灌注甘氨酰-谷氨酰胺(Gly-Gln)較甘氨酸可顯著提高肝臟移植鼠的回盲腸SIgA含量,增強腸黏膜屏障的功能[25]。
Gln在短腸綜合征的研究中發現,飼糧4%的Gln顯著提高小鼠糞中SIgA含量。試驗通過切除60%小腸、盲腸和1 cm結腸構建短腸模型(RX),對照組則在距回盲瓣60 cm處的小腸做橫切(TX),TX飼喂對照飼糧(TX/CON組),RX分別飼喂對照飼糧(RX/CON組)、4%Gln飼糧(RX/GLN組)和抗生素飼糧(RX/ABX組)。分別于手術前 1天,術后第 6、13、20天收集糞便,通過ELISA試劑盒檢測SIgA的含量,結果發現試驗第13天時,RX/GLN和RX/CON組SIgA含量顯著高于 RX/ABX和 TX/CON組,試驗第20天時,RX/GLN組SIgA含量顯著高于其余各組[26]。
為了確定Gln對SIgA分泌的影響是通過黏膜漿細胞產生的,該研究進行了免疫組織化學試驗,結果發現短腸鼠飼喂Gln飼糧(RX/CLN組)后較其他組顯著提高了殘余空腸固有膜IgA漿細胞的數量(圖1)[26]。進一步的研究還發現 RX/GLN組飼糧較RX/CON組對照飼糧降低細菌在腸道內的遷徙定植,而有促進小鼠增重的趨勢。
由此說明Gln在短腸綜合征中可促進SIgA的分泌,進而阻止病原菌在腸道的附著,有利于動物的生產性能。
4.3 維生素
以無菌SD小鼠為試驗動物,設4個組,分別是飼喂維生素A缺乏和正常飼糧組,以及飼喂維生素A缺乏和正常飼糧后再分別口服接種沙門氏菌組,接種后,待鼠糞便檢測到沙門氏菌時(4 d)屠宰小鼠,結果發現維生素A顯著影響腸黏膜SIgA含量,維生素A缺乏組(1.70 μg/mg)顯著低于維生素A正常組(2.75 μg/mg),而接種沙門氏菌后,SIgA的含量仍然是維生素A正常組(1.04 μg/mg)顯著高于維生素 A缺乏組(0.64 μg/mg)。且攻毒情況下維生素 A對腸道黏膜完整性表現出一定的保護作用(圖2)[27]。SIgA可以阻止微生物或其毒素產物在腸腔上皮細胞的附著從而呈現保護作用。
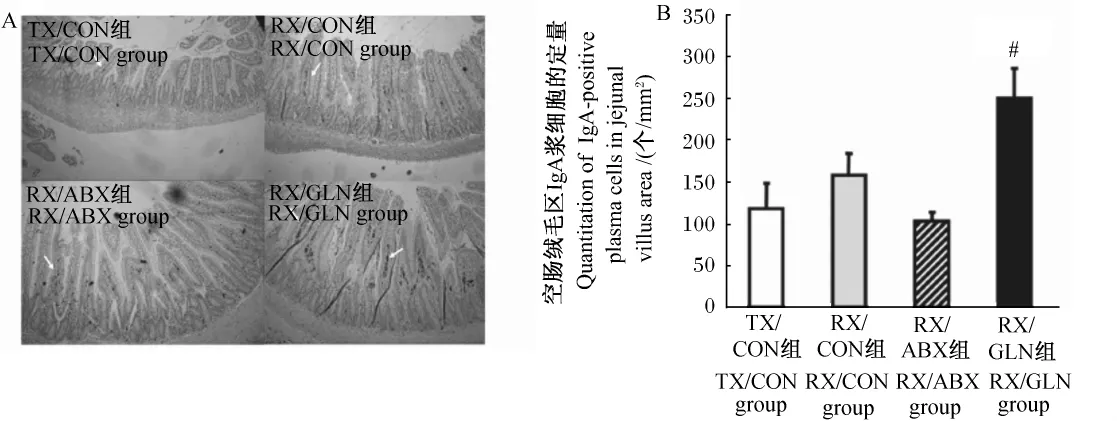
圖1 空腸中含IgA的漿細胞Fig.1 IgA-positive plasma cells in jejunum
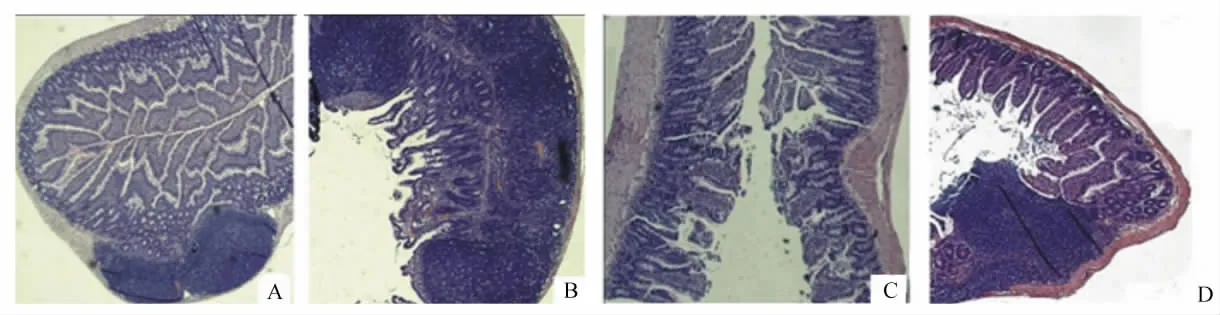
圖2 腸道黏膜組織病理學變化Fig.2 Histopathological changes of intestinal mucosa
白細胞介素 5受體 α(IL-5Rα)雜合型(IL-5Rα-/+)小鼠每天每只補充1 mg的維生素A 較 IL-5Rα 基因敲除型(IL-5Rα - /-)小鼠顯著提高黏膜IgA的含量及IgA漿細胞數量,且IgA的含量與維生素A呈現出劑量效應。當接種CT時,維生素A在IL-5Rα-/-基因敲除組未體現出促進SIgA分泌的效果,小鼠死亡率提高,而野生型(IL-5Rα+/+)鼠維生素A添加組顯著提高了腸道SIgA的含量并很好地抵御了CT的入侵,提高了小鼠存活率[28]。
4.4 其他營養因子
Otani等[29]研究發現,酪蛋白磷酸肽具有提高腸道SIgA含量和促進斷奶仔豬黏膜免疫的作用,研究發現酪蛋白磷酸肽一方面和靜止B細胞結合使其被激活;另一方面和T輔助細胞2(Th2)結合促進白細胞介素-5(IL-5)和白細胞介素-6(IL-6)的分泌,而活化的B細胞在增殖和分化IgA漿細胞過程中IL-5和IL-6起著重要作用。酪蛋白磷酸肽通過和Th2細胞結合促進 IL-5和 IL-6的分泌進而促進IgA的生成。項明潔等[30]在白血病患兒的研究中發現,處于白血病緩解期的患兒服用乳果糖后腸道中雙歧桿菌數量較服用前極顯著增加,SIgA含量顯著增加,腸道局部免疫力提高。另有研究發現,250、500 μg/g的海帶多糖能夠顯著促進小鼠腸黏膜SIgA的分泌量,進而提高小鼠的免疫力[31]。
以上研究表明,營養因子(如脂肪酸、氨基酸、維生素、酪蛋白磷酸肽、果糖等)可顯著促進機體SIgA的分泌,特別是在受到病原菌入侵或應激的情況下,這種改善作用尤為明顯,SIgA分泌的提高不僅有效地促進了機體對病原菌的抵抗,在一定程度上也可以改善應激動物的生產性能。但這些營養因子提高SIgA分泌,促進動物腸道免疫力增強的機制還有待深入研究。
5 小結
目前研究發現,SIgA在腸道黏膜免疫中通過維持腸道黏膜內環境穩態,干擾病原體與上皮細胞受體的結合,免疫排異清除病原體和降低病菌毒力,從而實現對機體的免疫保護,SIgA除以上作用方式外,是否還存在其他方式有效地提高機體免疫力仍需進一步研究。
已有研究發現,營養因子(如脂肪酸、氨基酸、維生素A等)在機體受到病原菌入侵或應激的情況下,顯著促進腸道SIgA的分泌,提高機體對病原菌的抵抗,然而這些研究大多集中在鼠上,其他動物上是否也存在類似的效果,以及其他營養因子能否促進腸道SIgA的分泌提高機體免疫力及其機制還有待探討。
[1]HENDRICKSON B A,RINDISBACHER L,CORTHéSY B.Lack of association of secretory component with IgA in J chain-deficient mice[J].The Journal of Immunology,1996,157(2):750 -754.
[2]VAERMAN J P,LANGENDRIES A,GIFFROY D,et al.Lack of SC/PigR-mediated epithelial transport of a human polymeric IgA devoid of J chain:in vitro and in vivo studies[J].Immunology,1998,95:90 - 96.
[3]JOHANSEN F E,BRAATHEN R,BRANDTZAEG P.Role of J chain in secretory immunoglobulin formation[J].Scand Journal of Immunology,2000,52(3):240-248.
[4]MAYER L.Mucosal immunity and gastrointestinal antigen processing[J].Journal of Pediatric Gastroenterology and Nutrition,2000,30(1):S4 - S12.
[5]EGAN C E,MAURER K J,COHEN S B,et al.Synergy between intraepithelial lymphocytes and lamina propria T cells drives intestinal inflammation during infection[J].Mucosal Immunology,2011,4(6):658-670.
[6]BRANDTZAEG P.Induction of secretory immunity and memory at mucosal surfaces[J].Vaccine,2007,25(30):5467-5484.
[7]FAGARASAN S,MURAMATSU M,SUZUKI K,et al.Critical roles of activation-induced cytidine deaminase in the homeostasis of gut flora[J].Science,2002,298:1424 -1427.
[8]REY J,GARIN N,SPERTINI F,et al.Targeting of secretory IgA to Peyer’s patch dendritic and T cells after transport by intestinal M cells[J].The Journal of Immunology,2004,172:3026 -3033.
[9]UREN T K,JOHANSEN F E,WIJBURG O L,et al.Role of the polymeric Ig receptor in mucosal B cell homeostasis[J].The Journal of Immunology,2003,170:2531-2539.
[10]APTER F M,LENCER W I,FINKELSTEIN R A,et al.Monoclonal immunoglobulin A antibodies directed against cholera toxin prevent the toxin-induced chloride secretory response and block toxin binding to intestinal epithelial cells in vitro[J].Infection and Immunity,1993,61:5271 -5278.
[11]APTER F M,MICHETTI P,WINNER L S D,et al.Analysis of the roles of antilipopolysaccharide and anti-cholera toxin immunoglobulin A(IgA)antibodies in protection against Vibrio cholerae and cholera toxin by use of monoclonal IgA antibodies in vivo[J].Infection and Immunity,1993,61:5279 -5285.
[12]HUTCHINGS A B,HELANDER A,SILVEY K J,et al.Secretory immunoglobulin A antibodies against the sigma 1 outer capsid protein of reovirus type 1 Lang prevent infection of mouse Peyer’s patches[J].The Journal of Immunology,2004,78:947 -957.
[13]HELANDER A,SILVEY K J,MANTIS N J,et al.The viral sigma 1 protein and glycoconjugates containing alpha 2-3-linked sialic acid are involved in type 1 reovirus adherence to M cell apical surfaces[J].Journal of Virology,2003,77:7964 -7977.
[14]MANTIS N J,FARRANT S A,MEHTA S.Oligosaccharide side chains on human secretory IgA serve as receptors for ricin[J].The Journal of Immunology,2004,172:6838 -6845.
[15]MESTECKY J,RUSSELL M W.Specific antibody activity,glycan heterogeneity and polyreactivity contribute to the protective activity of S-IgA at mucosal surfaces[J].Immunology Letters,2009,124:57 - 62.
[16]PERRIER C,SPRENGER N,CORTHESY B.Glycans on secretory component participate in innate protection against mucosal pathogens[J].Journal of Biological Chemistry,2006,281:14280 -14287.
[17]BRANDTZAEG P.Role of secretory antibodies in the defence against infections[J].International Journal of Medical Microbiology,2003,293:3 -15.
[18]LIéVIN-LE MOAL V,SERVIN A L.The front line of enteric host defense against unwelcome intrusion of harmful microorganisms:mucins,antimicrobial peptides,and microbiota[J].Clinical Microbiology Review,2006,19:315 -337.
[19]GALAN J E,WOLF-WATZ H.Protein delivery into eukaryotic cells by type Ⅲ secretion machines[J].Nature,2006,444:567 -573.
[20]RAFFATELLU M,SANTOS R L,VERHOEVEN D E,et al.Simian immunodeficiency virus-induced mucosal interleukin-17 deficiency promotes Salmonella dissemination from the gut[J].Nature Medicine,2008,14:421 -428.
[21]FORBES S J,ESCHMANN M,MANTIS N J.Inhibition of Salmonella enteric serovar typhimurium motility and entry into epithelial cells by a protective antilipopolysaccharide monoclonal immunoglobulin A antibody[J].Infect Immunology,2008,76:4137 - 4144.
[22]MANTIS N J,FORBES S J.Secretory IgA:arresting microbial pathogens at epithelial borders[J].Immunological Investigations,2010,39:383 -406.
[23]PéREZ-CANO F J,RAMíREZ-SANTANA C,MOLERO-LUíS M,et al.Mucosal IgA increase in rats by continuous CLA feeding during suckling and early infancy[J].Journal of Lipid Research,2009,50(3):467 -476.
[24]IMAEDA H,MIURA S,SERIZAWA H,et al.Influence of fatty acid absorption on bidirectional release of immunoglobulin A into intestinal lumen and intestinal lymph in rats[J].Immunology Letters,1993,38(3):253-258.
[25]JIANG J W,REN Z G,CHEN L Y,et al.Enteral supplementation with glycyl-glutamine improves intestinal barrier function after liver transplantation in rats[J].Hepatobiliary& Pancreatic Diseases International,2011,10:380 -385.
[26]TIAN J,LI H,PRAKASH C,et al.Dietary glutamine and oral antibiotics each improve indexes of gut barrier function in rat short bowel syndrome[J].American Journal of Physiology:Gastrointestinal and Liver Physiology,2009,296:G348 - G355.
[27]YANG Y,YUAN Y,TAO Y,et al.Effects of vitamin A deficiency on mucosal immunity and response to intestinal infection in rats[J].Nutrition,2011,27:227-232.
[28]NIKAWA T,IKEMOTO M,KANO M,et al.Impaired vitamin A-mediated mucosal IgA response in IL-5 receptor-knockout mice[J].Biochemical Biophysical Research Communication,2001,285(2):546-549.
[29]OTANI H,KIHARA Y,PARK M.The immunoenhancing property of a dietary casein phosphopetides preparation in mice[J].Food and Agriculture Immunology,2000,12(2):165 -173.
[30]項明潔,劉明,李云珠,等.乳果糖對腸道菌群及SI-gA含量的影響[J].江西醫學檢驗,2002,20(4):72-73.
[31]王庭祥,王庭欣,劉崢顥,等.海帶多糖對小鼠腸黏膜組織SIgA 的影響[J].安徽農業科學,2009,37(12):5515,5520.
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
