人結直腸癌耐藥細胞株HT-29/CPT-11的構建及其生物學特性探討
2012-09-03 01:49:38馮婉婷車曉玲李進宗明珠史玉葉曹維克何敬東
東南大學學報(醫學版) 2012年1期
關鍵詞:耐藥
馮婉婷,車曉玲,李進,宗明珠,史玉葉,曹維克,何敬東
(1.南京醫科大學,江蘇南京 210029;2.南京醫科大學附屬淮安第一人民醫院 腫瘤內科,江蘇淮安 223300)
結直腸癌(colorectal cancer,CRC)是消化道最常見惡性腫瘤之一,占所有惡性腫瘤的10% ~15%[1]。伊立替康(CPT-11)作為拓撲異構酶抑制劑家族成員之一,可抑制DNA單鏈斷裂后修復,干擾DNA復制和轉錄,導致腫瘤細胞死亡[2],現已廣泛應用于轉移性CRC的治療[3],無論是用于一線治療,還是在氟尿嘧啶治療失敗后的二線治療,CPT-11均顯示了其抗腫瘤的活性。然而化療藥物引起的耐藥是制約其療效的重要因素之一,因此,不斷提高患者化療敏感性是臨床化療亟需解決的重要課題。目前,有關預測CPT-11耐藥性的相關分子標志物的研究相對較少,主要集中在藥物代謝過程中參與的酶、轉運蛋白以及作用靶點蛋白的基因。人結腸癌耐藥細胞株的建立可為研究腫瘤耐藥機制、逆轉CRC耐藥提供實驗基礎。因此,本研究將采用逐步提高培養基中CPT-11濃度的方法,間歇誘導CRC細胞耐藥,并對親本細胞與耐藥細胞進行比較,初步探討其生物學特性及其耐藥機制。
1 材料與方法
1.1 材料
1.1.1 細胞株 人源性結直腸癌細胞株HT-29購自中國科學院上海生命科學研究所。
1.1.2 試劑與載體 CPT-11(Sigma公司),細胞周期檢測試劑盒(Beckman公司),RNA提取試劑Trizol(Invitrogen公司),逆轉錄及擴增試劑盒(Biouniquer公司),CCK-8細胞增殖檢測試劑盒(日本同仁化學研究所),1640培養基(美國Hyclone公司),胎牛血清(杭州四季青公司),PCR引物由上海生工生物工程有限公司合成。
1.2 方法
1.2.1 細胞培養及耐藥細胞的建立 將人結直腸癌細胞株HT-29置于含有10%胎牛血清、100 U·ml-1青霉素和100 μg·ml-1鏈霉素的 1640 培養基,置 37 ℃、5%CO2孵箱中培養。采用逐步提高培養基中CPT-11濃度的方法,間歇誘導腫瘤細胞耐藥。CPT-11濃度從4 μg·ml-1開始,加藥 48 h 后換液撤藥,3 ~4 d 換液1次,待細胞重新長至對數生長期時再次加相同濃度藥物進行誘導,如此重復再增加藥物濃度,連續培養10 個月,獲得能在 60 μg·ml-1濃度中穩定生長的人結直腸癌耐藥細胞株,并命名為HT-29/CPT-11。
1.2.2 CCK-8法檢測細胞藥敏性 取對數生長期的親本細胞HT-29和耐藥細胞HT-29/CPT-11,胰酶消化后,按每孔100 μl含2×104個細胞接種于96孔板上,培養24 h后分別加入藥物濃度分別為20、40、80、160、240、320 μg·ml-1的 CPT-11,設置 6 個復孔,培養48 h后,每孔加入CCK-8溶液10 μl,繼續培養2 h后,用全自動酶標儀在450 nm處測定OD值。計算藥物CPT-11分別對HT-29、HT-29/CPT-11的半數抑制率(IC50)。
細胞抑制率(%)=(1-實驗組平均OD值/對照組平均OD值)×100%。
耐藥指數(resistance index,RI)=子代細胞IC50/親代細胞IC50。
IC50采用改良寇氏法計算:Ig(IC50)=Xm-I(P-(3-Pm-Pn)/4)(Xm:Ig最大劑量,I:Ig最大劑量/相臨劑量,P:陽性反應率之和,Pm:最大陽性反應率,Pn:最小陽性反應率)
1.2.3 測定細胞生長曲線及倍增時間 將對數生長期的HT-29和HT-29/CPT-11細胞,以濃度為2×104ml-1分別接種在96孔板,間隔24 h分別取3個復孔,計數每孔活細胞數量,連續計數8 d,按每天計數細胞數量繪制細胞生長曲線。計算細胞的倍增時間公式:TD=t×log 2/(logNt-logNo)(其中TD:群體倍增時間,t:培養時間,N0、Nt分別代表接種后及培養t小時后的細胞數)。
1.2.4 流式細胞術(FCM)檢測細胞周期 收集處于對數生長期的HT-29和HT-29/CPT-11細胞,每組細胞數約1 ×106個,以1 500 r·min-1離心 5 min,PBS 洗2遍,預冷70%乙醇固定,再用PBS洗2遍,棄上清,在細胞沉淀中加入0.1%RNA酶A溶液150 μl重懸細胞,4 ℃ 孵育 30 min,再加入 0.1%PI染液 150 μl混勻,4℃避光孵育10 min,將細胞懸液混勻,用200目尼龍膜濾過到流式管中,加入1 ml PBS用FCM進行細胞周期分析,重復實驗3次。
1.2.5 RT-PCR半定量測定MDR-1 mRNA的表達分別收集處于對數生長期的HT-29和HT-29/CPT-11細胞,TRIzol一步法提取總RNA,按逆轉錄試劑盒說明操作(反應體系20 μl),合成第一鏈cDNA,擴增如試劑盒說明(反應體系25 μl)。引物序列:MDR-1上游引物5'-TCGTAGGAGTATCCGTGGAT-3',下游引物5'-CATTGGGCGAGCCTGGTAG-3';內參β-actin上游引物 5'-GCCTGGAAGTGAAGTTGTGGACTCCCG-3',下游引物 5'-CCAGCGTGAGTACTGCTGCGGCTCAG-3'。PCR循環條件:95℃ 3 min;94 ℃ 30 s,58.3℃ 30 s,72 ℃1 min,36個循環;72℃ 5 min。以β-actin作為內參,1%瓊脂糖凝膠100 V等壓電泳觀察實驗結果,凝膠成像儀掃描,以目的基因與β-actin光密度積分值之比作為其相對表達量(Image J)。
1.3 統計學處理
2 結 果
2.1 細胞形態學觀察
光鏡下觀察,發現親本細胞株HT-29呈圓形或橢圓形,細胞鑲嵌排列成鋪路石樣且成團生長(圖1)。耐藥細胞株HT-29/CPT-11比親本HT-29細胞體積明顯增大,形態也發生變化,排列紊亂無規律,細胞生長彌散。
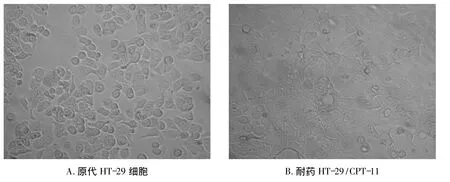
圖1 光鏡觀察細胞形態 ×10Fig 1 Light microscopy observation of cell morphology×10
2.2 藥物敏感性試驗
CCK-8檢測結果顯示,CPT-11對HT-29和HT-29/CPT-11 細胞的 IC50分別為(40.59 ± 3.29)μg·ml-1和(264.43 ± 8.98)μg·ml-1,耐藥指數為 6.51(圖2)。提示HT-29/CPT-11細胞對CPT-11有明顯的耐藥性(P<0.05)。
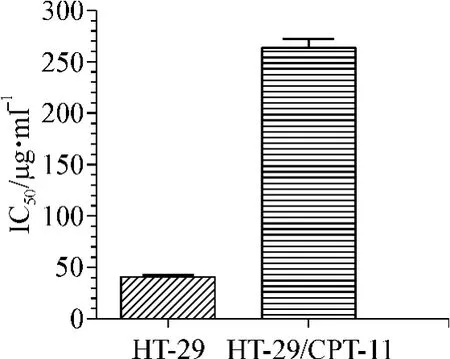
圖2 CPT-11對HT-29或HT-29/CPT-11的IC50Fig 2 IC50of CPT-11 in HT-29 or HT-29/CPT-11 cell lines
2.3 細胞生長曲線和群體倍增時間
細胞生長曲線(圖3)顯示,兩種細胞在6~7 d達到對數生長期,耐藥細胞株HT-29/CPT-11較HT-29群體倍增時間稍延長,分別為(41.29±1.69)h和(27.12 ±2.73)h,生長增殖速度減慢,兩者差異有統計學意義(P<0.05)。
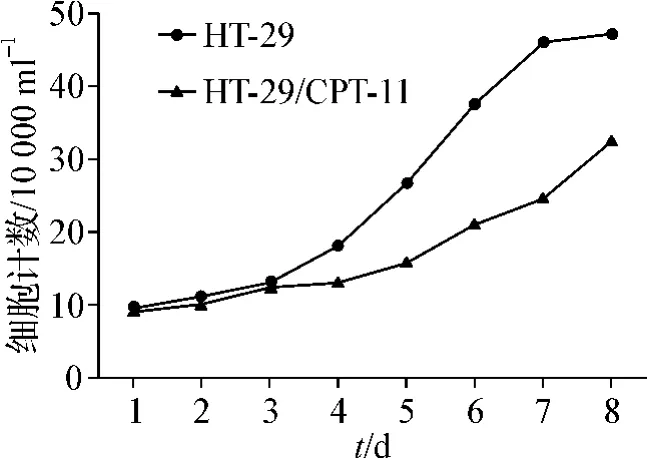
圖3 細胞生長曲線Fig 3 Growth curve of two cell lines
2.4 耐藥細胞株細胞周期的變化
見表1。
表1 HT-29和HT-29/CPT-11細胞周期比較(±s)Tab 1 Comparison of cell cycle between HT-29 and HT-29/CPT-11(±s)

表1 HT-29和HT-29/CPT-11細胞周期比較(±s)Tab 1 Comparison of cell cycle between HT-29 and HT-29/CPT-11(±s)
與 HT-29細胞比,a P <0.05
細胞株 G0/G1期 S期 G2/M期HT-2949.8 ±1.83 40.5 ±1.4 9.7 ±1.25 HT-29/CPT-11 60.2 ±2.71a 27.1 ±2.3a 12.7 ±1.8
由表1可見,與親本細胞相比,HT-29/CPT-11耐藥細胞的細胞周期分布發生明顯改變,G0/G1期細胞增加,S期細胞減少,差異有統計學意義(P<0.05)。
2.5 RT-PCR檢測MDR-1 mRNA表達
見圖4。
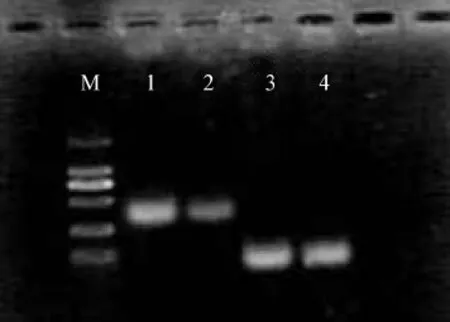
圖4 HT-29和HT-29/CPT-11細胞MDR-1 mRNA的表達Fig 4 The expression level of MDR-1 mRNA in HT-29 and HT-29/CPT-11 cell lines
Image J圖像分析結果顯示,HT-29/CPT-11、HT-29兩株細胞MDR-1/β-actin的光密度積分值之比分別為1.086 ±0.054 和 0.416 ±0.02(P<0.05),HT-29/CPT-11耐藥細胞MDR-1表達明顯高于親本細胞株HT-29,說明在CPT-11的長期刺激下可以誘導耐藥細胞MDR-1基因表達增強。
3 討 論
細胞耐藥既是正常細胞維持自身穩定的防御機制之一,也是引起腫瘤化療失敗及腫瘤復發的主要原因之一。腫瘤細胞耐藥機制十分復雜,迄今為止,體外誘導耐藥細胞株仍是研究細胞分子生物學變化及腫瘤耐藥機制的基礎。
近年來,國內外相繼建立了胃癌、乳腺癌、卵巢癌等耐藥細胞模型。我們采用逐步提高培養基中CPT-11濃度的方法,間歇誘導CRC耐藥細胞株,歷經10月余,成功建立并鑒定了人結直腸癌CPT-11耐藥細胞株HT-29/CPT-11。其對 CPT-11耐藥指數為6.51。且耐藥細胞株HT-29/CPT-11細胞形態異型性明顯增加。除此之外,細胞倍增時間顯著延長。目前認為,腫瘤細胞倍增時間越短,對化療藥物越敏感,療效越好;反之,對化療藥物的敏感性降低。流式結果顯示G0/G1期細胞增加,S期細胞減少,說明細胞在耐藥之后其增殖受到明顯抑制,DNA復制時間延長。
腫瘤的多藥耐藥(multidrug resistance,MDR)機制是一個復雜的生物學過程,影響因素和參與機制眾多。目前,關于腫瘤耐藥機制的研究主要集中以下幾點:(1)轉運蛋白引起的耐藥,包括有腫瘤MDR基因[4]編碼產物的P-糖蛋白(p-glycoprotein,p-gp)的高表達、MDR相關蛋白(multidrug resistance associated protein,MRP)基因及其產物的表達[5]。(2)GSH 依賴性解毒酶系統的改變[6],谷胱甘肽 S轉移酶(ghtathione S-transferase,GST)參與化療藥物的解毒,降低抗腫瘤藥物的細胞毒作用,從而產生耐藥。(3)當DNA損傷修復時,參與修復的相關酶,包括核酸內切酶和DNA連接酶等數種酶活性增強。細胞修復損傷的能力和MDR產生密切相關[7]。(4)細胞外低pH值、細胞質內高pH值的特點,不僅可造成化學治療藥物被隔離在酸性區內,還能上調P-gp表達和活性,結果使化學治療藥物無法到達細胞內靶點發揮作用,導致腫瘤對化療藥物耐藥[8]。
細胞內藥物蓄積減少是最先發現的、也是最常見的導致腫瘤耐藥的原因之一。有研究[5-9]顯示,參與藥酶代謝的ABC轉運蛋白家族可以減少細胞之間藥物的蓄積,從而使得細胞對喜樹堿類藥物耐藥。膜轉運蛋白中P-gp、MRP2和BCRP是參與CPT-11代謝最重要的轉運蛋白。耐藥細胞表面P-gp的發現使人們認識到藥物泵對藥物的主動外排是細胞內藥物蓄積減少的主要原因[5]。MDR-1基因的過度表達是產生多藥耐藥的主要機制,這一觀點已得到公認[4]。MDR-1基因編碼跨膜糖蛋白P-gp,P-gp具有“藥泵功能”,是一種ATP依賴性藥泵,通過分解ATP提供能量,將藥物轉運到細胞外,使細胞內藥物濃度維持較低水平,從而產生耐藥。腫瘤細胞在長期的化療藥物的刺激下,耐藥基因及其編碼蛋白表達的改變可以進一步證實細胞株的耐藥性。因此,本實驗將MDR-1基因作為研究對象,采用RT-PCR技術檢測耐藥細胞株耐藥細胞株HT-29/CPT-11和親本細胞株HT-29中MDR-1基因的表達水平,發現耐藥細胞株MDR-1基因表達明顯增加(1.086 ±0.054vs0.416 ±0.02,P<0.05),這說明在CPT-11的長期刺激下可以誘導耐藥細胞MDR-1基因表達增強,提示MDR-1基因的高表達可能是造成HT-29/CPT-11細胞內CPT-11藥物蓄積減少,從而成為耐藥發生的可能機制之一。當然除了細胞MDR-1基因參與之外,還有其他途徑參與,有待進一步研究。
總之,本研究結果表明,我們利用藥物濃度遞增間歇誘導出的人結直腸癌CPT-11耐藥細胞株在細胞形態、細胞周期、生長速度上以及藥物敏感性等方面發生了明顯的變化,符合耐藥細胞的特征。在下一步研究中,我們將利用耐藥細胞株HT-29/CPT-11進行耐藥機制以及耐藥逆轉方面的研究。
[1]JEMAL A,SIEGEL R,WARD E,et al.Cancer statistics[J].CA Cancer J Clin,2008,58(2):71-96.
[2]INNOCENTI F,KROETZ D L,SCHUETZ E,et al.Comprehensive pharmacogenetic analysis of irinotecan neutropenia and pharmacokinetics[J].J Clin Oncol,2009,27(16):2604-2614.
[3]ROUGIER P,MITRY E.Colorectal cancer chemotherapy:irinotecan[J].Semin Oncol,2000,27:138-143.
[4] GILLET J P,EFFERTH T,REMACLE J.Chemotherapy-induced resistance by ATP-binding cassette transporter genes[J].Biochim Biophys Acta,2007,1775(2):237-262.
[5]ECKFORD P D W,SHAROM F J.ABC efflux pump based resistance to chemotherapy drugs[J].Chem Rev,2009,109:2989-3011.
[6]LO H W,ALI-OSMAN F.Genetic polymorphism and function of glutathione S-transferases in tumor drug resistance [J].Curr Opin Pharmacol,2007,7(4):367-374.
[7]FERGUSON D O,ALT F W.DNA double strand break repair and chromosomal translocation:lessons from animal models[J].Oncogene,2001,20(40):5572-5579.
[8]HARGUINDEY S,ORIVE G,LUIS PEDRAZ J,et al.The role of pH dynamics and the Na+/H+antiporter in the etiopathogenesis and treatment of cancer.Two faces of the same coin-one single nature[J].Biochim Biophys Acta,2005,1756(1):1-24.
[9]YANG C H,SCHNEIDER E,KUO M L,et al.BCRP/MXR/ABCP expression in topotecan-resistant human breast carcinoma cells[J].Biochem Pharmacol,2000,60(6):831-837.
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
