渤海灣西南部典型站位營養鹽限制特性的加富培養實驗研究
2012-08-14 05:48:52穆迪李清雪陶建華趙海萍聶紅濤
海洋通報 2012年6期
穆迪,李清雪,陶建華,趙海萍,聶紅濤
(1.河北工業大學 海水資源高效利用化工技術教育部工程研究中心,天津 300130;2.河北工程大學 分析測試中心,河北 邯鄲 056038;3.天津大學 環境科學與工程學院,天津 300072)
近年來,隨著一些海域富營養化和赤潮問題的加劇,以及利用海洋生物泵吸收CO2緩解溫室效應問題的升溫,營養鹽對于浮游植物的限制作用再次成為海洋學研究的熱點(劉慧等,2002)。加富培養實驗直觀、可操作性強,接近于自然狀況,是研究營養鹽限制特性的常用方法之一(蒲新明等,2000)。在我國,這項研究始于20世紀末,研究海域主要集中在黃渤海、東海近海海域(鄒立等,2001;王勇等,1999;劉媛等,2004)、臺灣海峽(王玉玨等,2008;胡俊等,2008)及膠州灣(王勇等,2002)、大亞灣(王曉偉等,2007)等海域,結果表明,氮和磷是這些海域主要的限制性營養鹽,但具體的限制類型又體現了較大的時空差異性。目前這類實驗的目的主要還是營養鹽限制類型的判定,對培養條件生態影響的研究則較少。氮磷比是影響浮游植物生長的重要因素之一(Hodgkiss et al,1997),各種浮游植物對氮磷營養鹽的需求不同,一個海域的最適宜氮磷比通常具有特異性,Redfield比只是一個平均水平(Hecky et al,1988),但以往的多因素培養實驗多是基于Redfield比進行設計,沒有對所研究海域的最適宜氮磷比進行定量分析并加以考慮。唐松(2010)在印度洋進行氮、磷、鐵加富的現場培養實驗時,分析了實驗過程中氮磷比與浮游植物生長之間的關系,認為二者相關性不大,氮磷比不能單獨決定浮游植物的生長,但該實驗是在相對獨立、加富條件差異較大的培養組間進行的,此時營養鹽的絕對濃度影響更大,對于氮磷比的作用還需在相對一致的條件下進行系統研究。此外,以往的加富培養實驗采用的多是營養鹽的一次性添加,這種添加方式更接近沖擊性高負荷排污的情況,并不適合常見的排污情況,對于不同添加方式的影響應予以考慮。雖然王玉玨等(2008)在進行臺灣海峽的研究時使用了連續添加,但是沒有對同等藥量的一次性添加進行對比,從而分析添加方式的影響。
渤海灣位于渤海西部,屬典型的緩坡淤泥質半封閉淺海海灣。渤海灣沿岸有海河、永定新河等幾十條河流注入,每年有大量污水入海。該海域水交換能力弱,污染物滯留時間長,致使該海域污染嚴重、赤潮頻發(Tao,2002)。目前,對于渤海灣內營養鹽限制及其相關特性的調查研究多是基于水體中溶解無機營養鹽的現存量進行的對比分析(Zou et al,1985;沈志良,1999;闞文靜等,2010)。這些研究表明,近年來渤海灣近岸海域水體營養鹽的比例和含量發生了較大變化,進一步研究營養鹽比例和含量的變化對浮游植物生長的影響是十分必要的。
本次研究的主要工作是通過改進實驗設計,對渤海灣西南部海域典型站位2010年10月的營養鹽限制特性進行加富培養的實驗研究。研究的主要內容包括營養鹽限制類型的判定,浮游植物對于不同氮磷比和不同硝酸鹽添加方式的生態響應。本次研究不僅是一次實驗調查,也是一次實驗改進的嘗試。
1 材料與方法
1.1 采樣方法
采樣站位位于渤海灣西南部(117°49′56″E,38°38′4″N),為渤海灣生態監控區常規監測站位之一,采樣時間為2010年10月。采樣時采集表層水,采樣后用200 μm的篩網輕柔過濾,以濾除大型浮游動物,避免攝食干擾。
1.2 實驗方法
水樣采集后,在常溫狀態下,用汽車迅速運回實驗室,進行混合、分裝和培養,水樣的采集和運輸共耗時6 h左右。每桶初始水量15 L,進行營養鹽添加后將桶置于光照培養架上進行培養。用空調對培養室室溫進行了調節,日間控制室溫在18℃~20℃左右,夜間利用常溫培養,室溫最低在11℃~13℃左右。光照強度控制在初始水位處光強6 000 lux,明暗周期為12-12 h。光照是影響浮游植物光合作用的重要因素,在海上進行現場培養實驗時通常采用自然光照(鄒立等,2001;王勇等,1999;王玉玨等,2008)或維持培養環境與現場接近(劉媛等,2004;胡俊等,2008;王勇等,2002),在室內進行模擬現場培養實驗時為模擬現場環境或控制適宜培養條件通常需要控制光照(韋桂秋,2006;鄧華健等,2004),本文光照強度的設置主要考慮將其控制在浮游植物生長的適宜范圍。一般而言,在一定范圍內,浮游植物的光合作用隨著光照的增強而增強,但達到一定強度(飽和光強)后,光強的增加不再具有促進作用,反而會對光合作用產生抑制,浮游植物最適宜的光強范圍一般在 2 000~10 000 lux(鄧華健 等,2004;孫霞,2005)。本文參照此規律及相關研究結果并考慮了水深的影響,選用了6 000 lux。12-12 h的明暗周期是比較常見的模擬實際光照的方法。
每天固定時間混合水樣3次,防止浮游植物聚集。培養周期為14 d。本文浮游植物培養周期的設置主要參考了相關實驗研究。文獻表明,加富培養實驗的培養周期通常在3~12 d不等(王曉偉等,2007;胡俊等,2008;鄧華健等,2004),或動態的指定到某一生長階段(如指數生長期/最大增值期第4 d)(韋桂秋,2006),本文考慮到實驗初期的生長延遲現象,最終確定培養周期為14 d。浮游植物培養周期的選取主要與浮游植物的生長周期有關,應盡量使二者相吻合,能在培養期間清楚地觀察到整周期的生長變化。
營養鹽添加時,硝酸鹽、磷酸鹽和硅酸鹽分別以KNO3、KH2PO4、Na2SiO3形式進行添加。采樣和實驗所用器具使用前均使用HCl漬洗和海水潤洗。
1.2.1 實驗一:營養鹽限制類型的加富培養實驗
當浮游植物受到某種營養鹽限制時,人為添加這種營養鹽,會促進浮游植物的生長。把一種和多種營養鹽加入到一定水體中,通過觀察比較浮游植物的生長情況,判斷其中的營養鹽限制情況,是用加富培養實驗判定營養鹽限制類型的原理和設計依據。這種判定方法也稱AGP(Algal Growth Potential,藻類增長潛力)法(韋桂秋,2006)。
無機氮(硝酸鹽、亞硝酸鹽、氨氮)、磷酸鹽和硅酸鹽是對海洋浮游植物生長最重要的幾種營養鹽。陸源排放是渤海灣營養鹽的主要來源,近年來渤海灣營養鹽陸源污染嚴重、比例失調,近岸海域水體營養鹽的比例和含量發生了較大變化(闞文靜等,2010),實驗一即為驗證水體營養鹽的比例和含量發生變化的情況下,浮游植物的營養鹽限制類型是否也發生變化。
本實驗中,為判定原水樣的營養鹽限制類型,對水樣進行了硝酸鹽、磷酸鹽、硅酸鹽的單一和全部添加(表1)。在浮游植物適宜范圍內,在系統本底濃度的基礎上按倍數增大營養鹽的濃度是加富培養實驗的一種常見的營養鹽添加方式,文獻中的增大倍數在2倍(鄒立等,2001)、10倍(劉媛等,2004;王曉偉等,2007)和100倍(王曉偉等,2007)不等。本文營養鹽的添加量也是依據營養鹽的本底濃度確定的。其中氮鹽增大為本地濃度的30倍左右,磷酸鹽增大為本底濃度的20倍左右,硅酸鹽增大為本底濃度的15倍左右,進行取整,并考慮了同時添加時各營養鹽濃度的比值接近Redfield比。
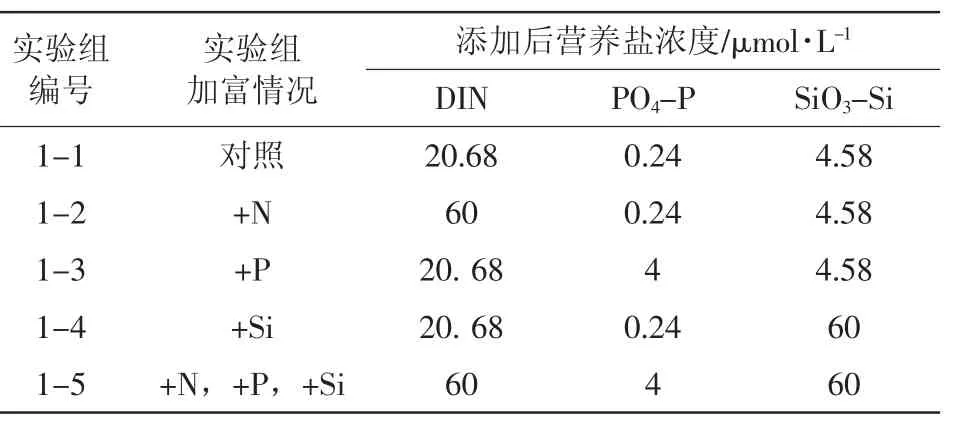
表1 實驗一加富方案
1.2.2 實驗二:氮磷比生態響應的加富培養實驗為考察不同氮磷比對于浮游植物生長的影響,在保證營養鹽絕對值充足的情況下(Fisher et al,1999;Nelson et al,1995),添加和控制硝酸鹽濃度為一定水平,變化磷酸鹽濃度,設計了8個實驗組(表2),代表8種不同的氮磷比水平。各實驗組濃度梯度的選取主要考慮了普遍條件下適宜浮游植物生長的氮磷比范疇和渤海灣常見的氮磷比范疇(蒲新明 等,2000;Hodgkiss et al,1997;Zou et al,1985;沈志良,1999;闞文靜等,2010),并考慮了添加和測定的精度要求。
1.2.3 實驗三:硝酸鹽添加方式生態響應的加富培養實驗
設計了4組對照實驗,考察硝酸鹽一次性添加和硝酸鹽連續性添加情況下,浮游植物生態響應的差異(表3)。其中,硝酸鹽一次性添加是模擬沖擊性高負荷排污情況,連續性添加是模擬持續性低負荷排污情況。這里,營養鹽的連續性添加是指,將與一次性添加等量的營養鹽均勻地分7天連續添加,每日藥量的計算中考慮了每日取樣量對體積的影響。這里控制磷酸鹽濃度一定,考察硝酸鹽添加的影響主要是基于近年來渤海灣氮污染嚴重的特點(闞文靜等,2010)。
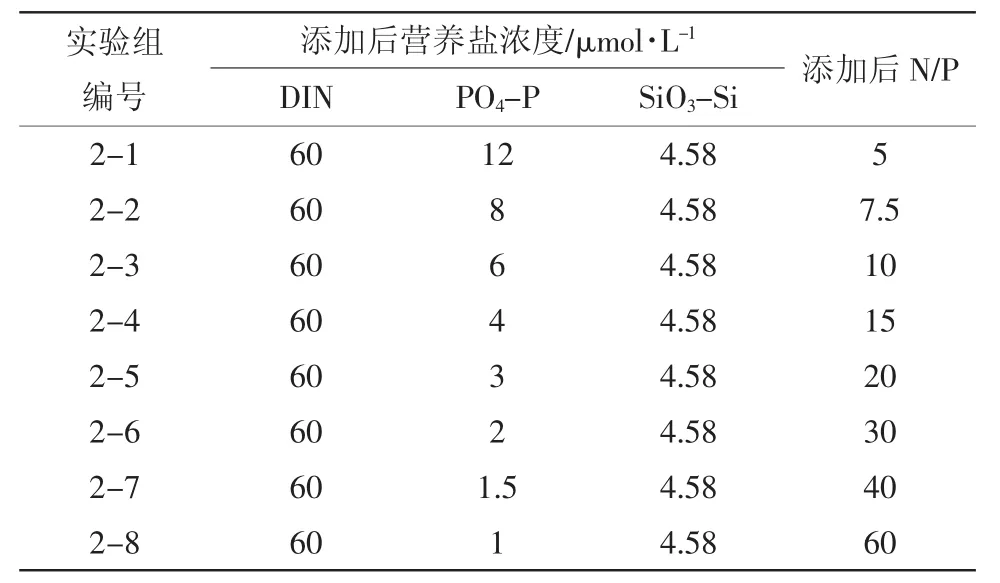
表2 實驗二加富方案
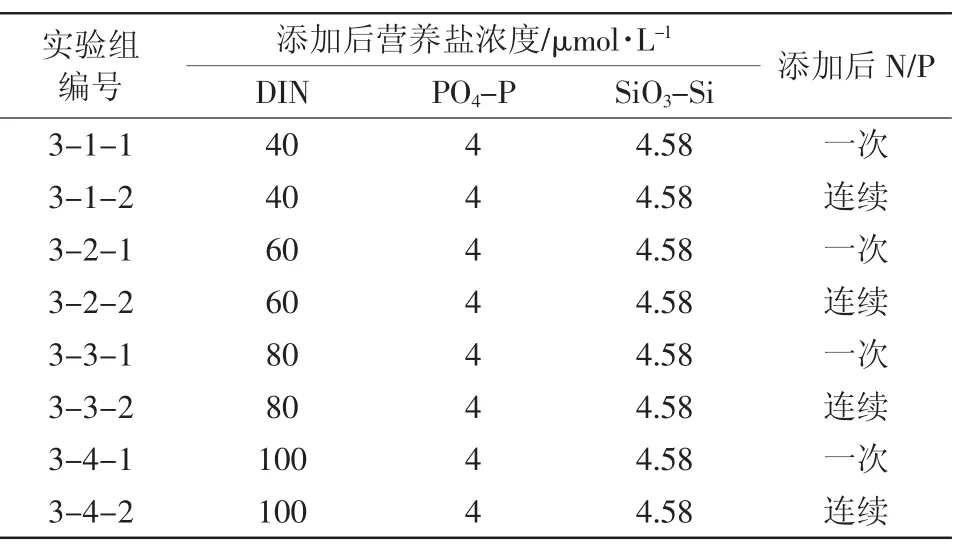
表3 實驗三加富方案
1.3 分析方法
2 結果與討論
2.1 原水樣水質
原水樣中溶解無機氮濃度為20.68 μmol/L,其中:硝酸鹽濃度為16.58 1 μmol/L,亞硝酸鹽濃度為 2.356 μmol/L,氨氮濃度為 1.746 μmol/L。水樣中磷酸鹽濃度0.24 μmol/L,硅酸鹽濃度4.58 μmol/L(表1)。原水樣中葉綠素a濃度為1.05 μg/L,浮游植物細胞密度為1 080 cells/L,主要有甲藻和硅藻,數量占優勢的藻種為叉狀角藻和園篩藻。在水樣運輸過程中,可能出現少量浮游植物死亡,釋放營養鹽的情況。
2.2 營養鹽限制類型
培養期間,單一添加營養鹽的各組中,添加磷酸鹽的1-3組葉綠素a濃度和浮游植物細胞密度的增長狀況最顯著,在培養的前8天時間里,1-3組葉綠素a濃度和浮游植物細胞密度的變化趨勢最接近于營養鹽全部添加的1-5組。其中,1-3組葉綠素a濃度峰值為空白對照組1-1組的2.48倍,達到營養鹽全加組1-5組同期濃度的48%,其細胞密度峰值為1-1組的1.66倍,達到1-5組同期密度的72%,這里只給出了細胞密度的變化趨勢(圖1(a))。同時,各組磷酸鹽都出現了明顯的消耗,在沒有磷酸鹽添加的情況下都是最早被耗盡(圖1(b))。硅酸鹽的消耗也較顯著,在沒有硅酸鹽添加的各組中,最終都趨于耗盡,耗盡時間僅晚于磷酸鹽(圖1(c))。培養期間,各組溶解無機氮濃度波動較大,總體變化不明顯。從浮游植物生長和營養鹽消耗的特征來看,該水樣為磷限制。
同時,對原水樣營養鹽濃度進行對比分析表明,若以Fisher(1999)或Nelson(1995)的閾值標準評定氮、磷、硅營養鹽都不構成限制,若以Justic(1995)和Dortch(1992)的比例關系評定,則該水樣接近于磷限制(Si∶P=19.08,DIN∶P=86.17),這與上述培養實驗結論基本一致。
綜合以往我國不同海域的實驗成果表明,氮和磷是這些海域主要的限制性營養鹽,營養鹽限制特性具有明顯的時空差異性(表4)。同時,由于近岸地化過程和人類活動的影響,近岸海域營養鹽限制特性通常呈現一定的復雜性(張平等,2001)。磷限制通常發生在沿岸和較封閉海域(劉慧等,2002),我國曾發現磷限制的海域包括渤海萊州灣附近(鄒立等,2001)、大亞灣的灣中和灣頂(王曉偉等,2007)和東海長江口附近站位(劉媛等,2004)。近年來,由于排污,一些近岸海域出現了營養鹽濃度增高比例失調,營養鹽限制特性發生轉變的情況。如對膠州灣進行的培養實驗(王勇等,2002)和營養鹽濃度分析(張均順等,1997)表明,膠州灣水體正在趨向于硅限制。渤海灣,屬典型的緩坡淤泥質半封閉淺海海灣,水交換能力弱,自凈能力差。近年來隨著沿岸經濟的快速發展,渤海灣營養鹽的含量和比例發生了顯著的變化,表層海水氮磷比和硅磷比增加,硅氮比較穩定,且小于1,近年來渤海灣浮游植物可能正在從氮限制逐漸轉變為磷限制或硅限制(闞文靜等,2010)。本文的實驗結果與此前報道基本一致。
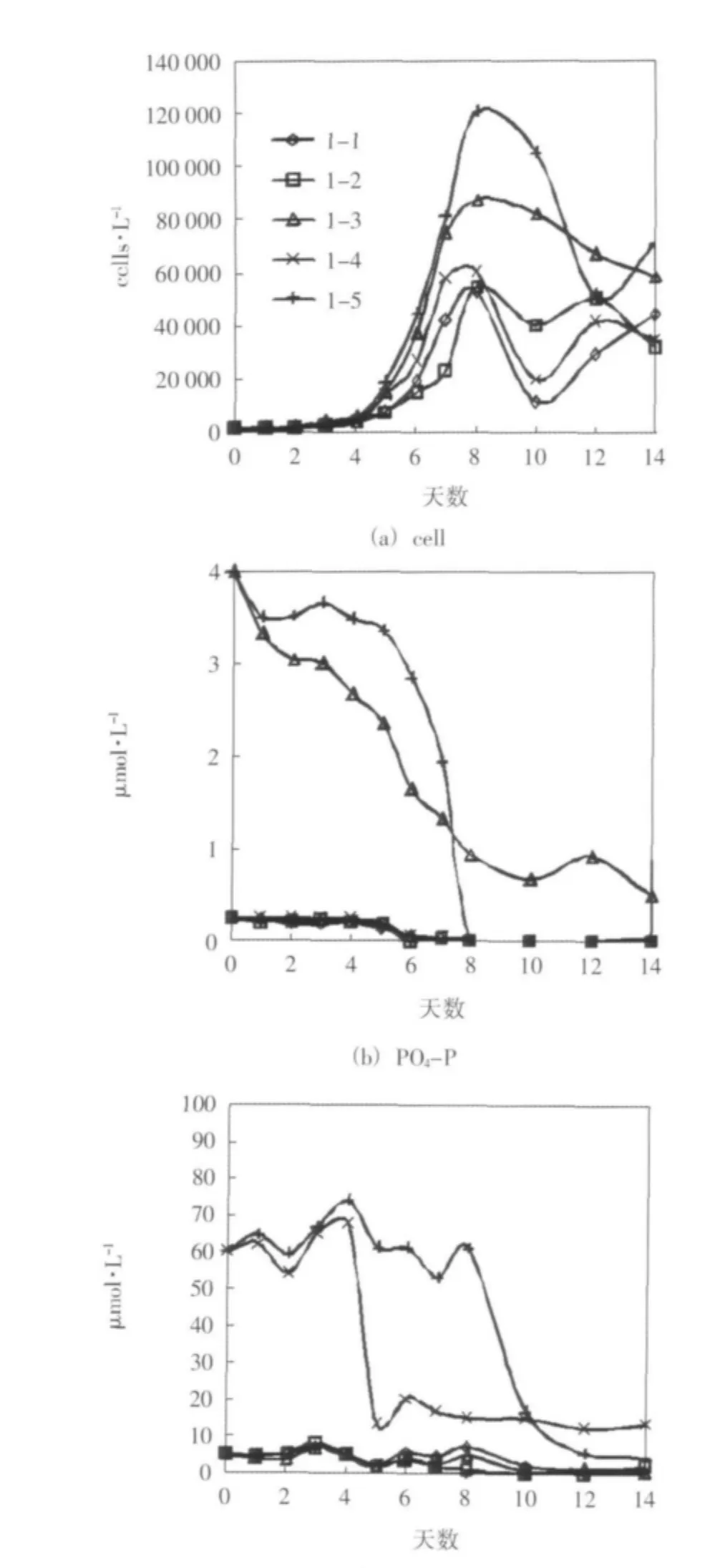
圖1 實驗一 培養期間細胞密度和營養鹽濃度的變化趨勢
2.3 不同氮磷比的生態響應
對各組在培養期間葉綠素a濃度、浮游植物細胞密度的變化趨勢進行了對比,并對前八天時間里不同氮磷比對應的葉綠素a濃度最大值和細胞密度最大值進行了統計。差異較顯著的前五組的細胞密度隨時間的變化的情況如圖2(a)所示(后3組的細胞密度總體水平較低,差異不顯著,這里沒有給出)。各組細胞密度最大值與對應氮磷比的關系如圖2(b)所示。
結果表明,葉綠素a濃度和浮游植物細胞密度的增長總體上隨磷酸鹽濃度的升高和氮磷比的降低而增大,并在5-15附近增長最顯著,這個范圍可以判定為該實驗條件下浮游植物生長的最適宜氮磷比,這個氮磷比的范圍與香港海域主要赤潮藻種的最適宜氮磷比4-16的范圍接近(Ho et al,1993)。相比于Redfield比,這個最佳氮磷比范圍偏低,這可能與樣品中甲藻較多有關,甲藻對磷的需求量更大,適宜低氮磷比的環境(蕭樸云等,2009)。另外,本次實驗是在不添加硅酸鹽、氮硅比較高(DIN∶DISi=13.1)的情況下進行的,可能存在硅的相對不足而限制硅藻的生長。根據本實驗可以推測,如果渤海灣磷酸鹽濃度增大,氮磷比降低,渤海灣的富營養化可能造成更為嚴重的后果,這從另一方面體現了磷的限制作用。
相對于以往的研究(唐松等,2010),本次實驗體現了較好的規律性,這可能與本實驗中控制無機氮、硅酸鹽初始濃度相同,系統條件相對一致有關,同時實驗中營養鹽濃度偏離正常值較多,這可能促進了少數耐污種類的生長,降低了浮游植物的多樣性,從而使響應更單一,這雖然與現實情況有所偏差,但是可以更清晰地反映浮游植物的響應規律。營養鹽變化對浮游植物種類的影響還有待進一步研究。
2.4 不同硝酸鹽添加方式的生態響應
對各對照組在培養期間葉綠素a濃度、浮游植物細胞密度和營養鹽的濃度變化進行了兩兩對比。生長較顯著的3-2組(3-2-1、3-2-2)的情況如圖3所示。其他3個對照組的變化規律基本相似,但沒有3-2組浮游植物的生長顯著、差異明顯,這里沒有給出。對照組間的這種差異可能與營養鹽條件不同有關,3-2組的磷酸鹽含量和氮磷比均適宜浮游植物的生長;3-1組雖然氮磷比適宜,但磷酸鹽含量偏低;3-3與3-4組無機氮源充足但氮磷比偏高,故3-2組浮游植物的生長更顯著。

表4 國內近岸海域加富培養實驗研究及其成果
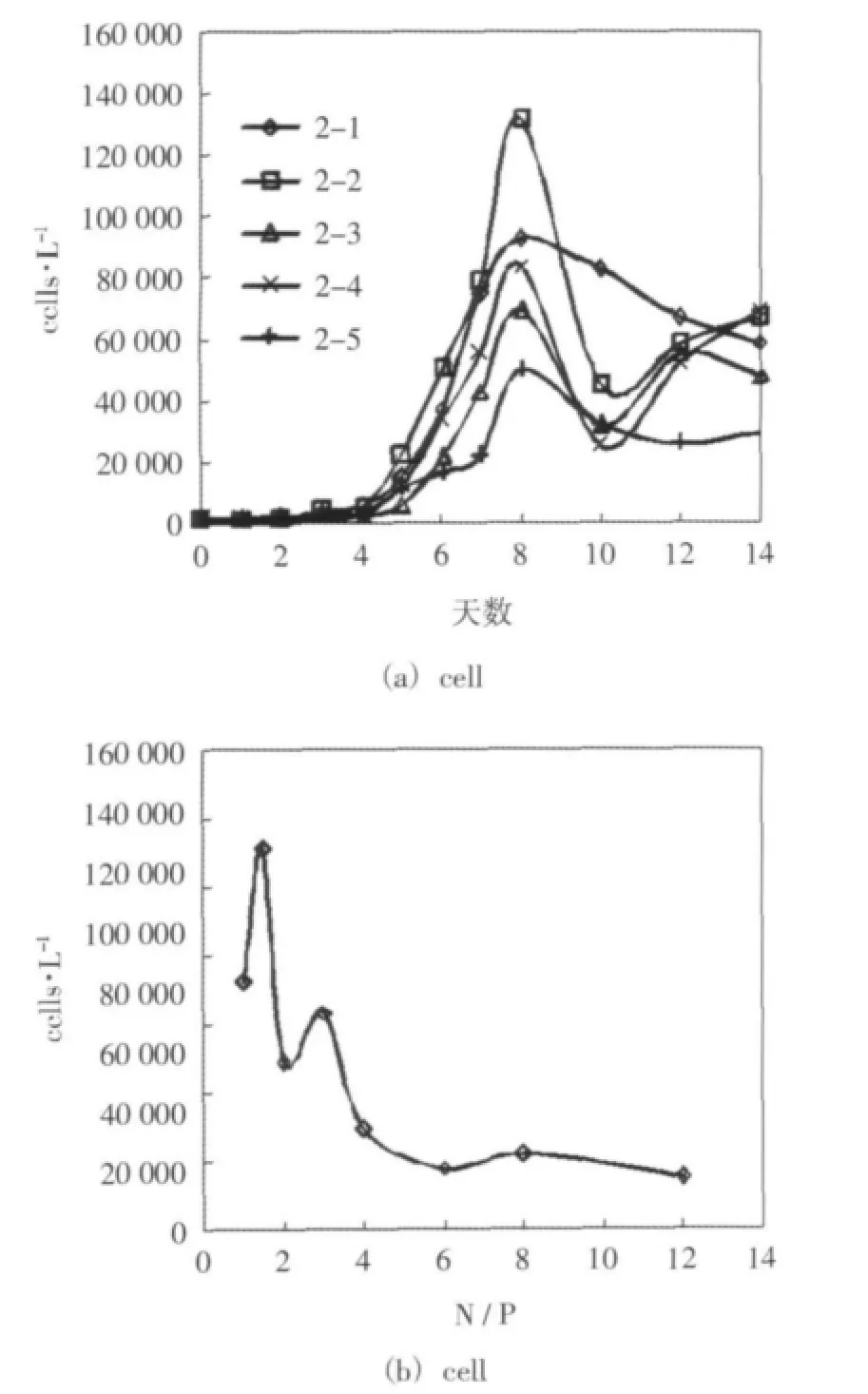
圖2 實驗二 培養期間細胞密度的變化趨勢和細胞密度最大值與氮磷比的關系
結果表明,在其他條件相同的情況下,硝酸鹽分七天連續添加時相對于一次性添加時能達到更高的葉綠素a濃度和浮游植物細胞密度。在浮游植物生長旺盛時期,連續性添加組的磷酸鹽較一次性添加組的磷酸鹽吸收快、濃度低。硝酸鹽濃度變化的波動較大,但前期連續性添加組濃度逐漸攀升的趨勢明顯。在培養前期,由于營養鹽添加方式的不同,連續性添加組的氮磷比低于同期一次性添加組的氮磷比,按上文給出的規律,這可能促進了連續性添加組磷的吸收和低氮磷比浮游植物的生長,但這種優勢隨著硝酸鹽的連續添加會逐漸縮小。同時,在培養的后期連續性添加組維持了較高的硝酸鹽濃度水平,這對于后期浮游植物的生長可能也是有利的。
該實驗結果表明低濃度長期持續性氮污染可能會比高濃度沖擊性氮污染更有效地刺激浮游植物的增長,從而造成更嚴重的生態影響。同時該結果也表明在本實驗條件下,用以往的一次性添加培養實驗可能會低估浮游植物的增長潛力。
2.5 與渤海灣海域實際狀況的比較
為分析自然狀況下該海域營養鹽變化時浮游植物的變化情況,對20世紀50年代末至21世紀初渤海灣水體營養鹽和浮游植物的幾次大規模調查研究進行了搜集整理(表5和表6)。綜合結果表明,半個世紀以來渤海灣氮磷營養鹽含量整體上呈現增長的趨勢,尤其是近年來溶解無機氮增長很快,而水體磷酸鹽濃度得到一定控制,結果造成水體氮磷比逐年增大,明顯偏離了Rieldfied比,水體基本處于磷限制。同期,渤海灣浮游植物的生態特性也發生了較大變化,浮游植物細胞豐度增大,多樣性降低,組成更單一,且時空分布極不均勻。水體營養鹽濃度和比例結構的變化可能是造成浮游植物群落結構變化的一個直接原因,而根本的原因還是受到陸源污染的影響。
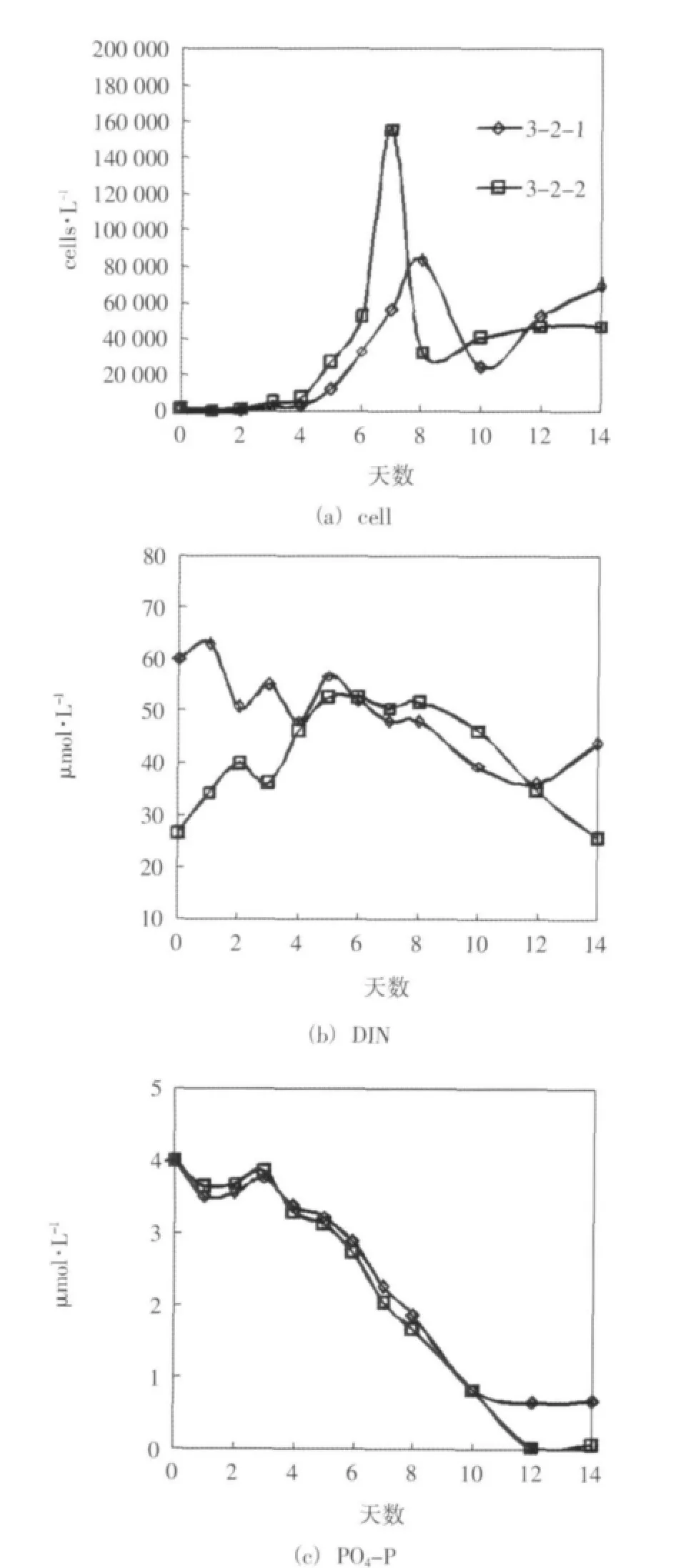
圖3 實驗三 培養期間細胞密度和營養鹽濃度的變化趨勢
本文在控制條件下,對渤海灣西南部海域典型站位表層水體進行了模擬現場的加富培養實驗,研究了水體的營養鹽限制類型以及不同氮磷比和不同硝酸鹽添加方式的生態響應。結果表明水體處于磷限制,這與海域的實際情況一致。同時,實驗結果表明,在保持氮充足的情況下變化磷酸鹽濃度時,較低的氮磷比(5-15)更能促進浮游植物的生長,根據該結論可以推測,如果渤海灣磷酸鹽濃度增大,氮磷比降低,渤海灣的富營養化可能造成更為嚴重的生態后果。近年來,伴隨著水體氮磷比增大,渤海灣浮游植物也出現了總量增大的趨勢,這主要還是受多種營養鹽,尤其是氮污染加劇的影響,與本文實驗條件的情況是不同的。對于不同排污方式實際影響的研究目前還未見報道,因此沒有進行對比分析。
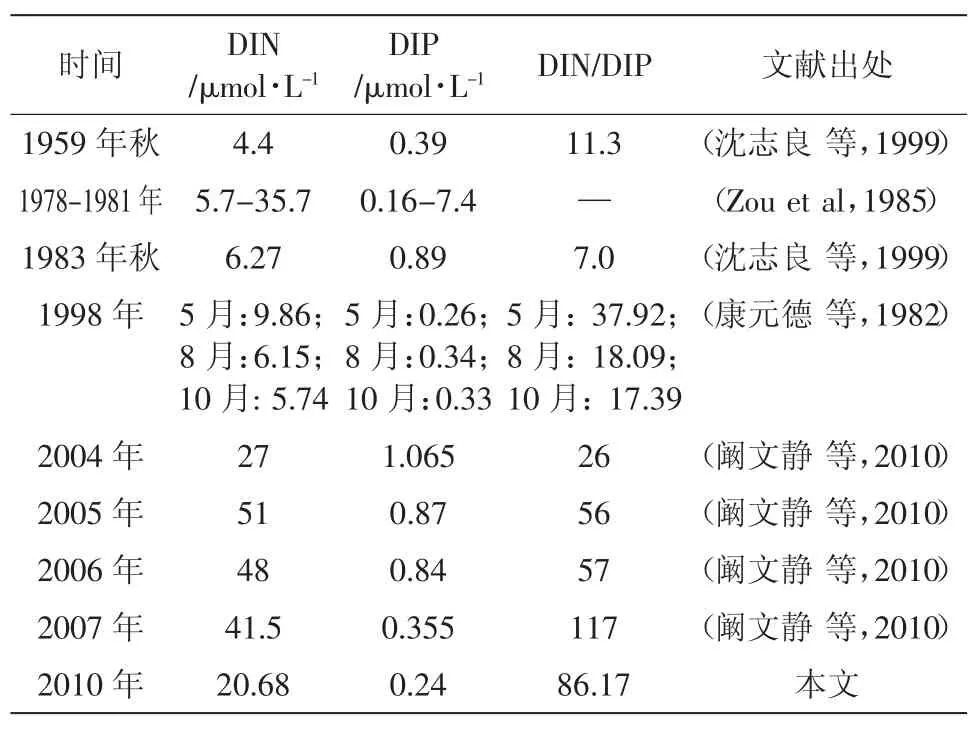
圖3 實驗三 培養期間細胞密度和營養鹽濃度的變化趨勢
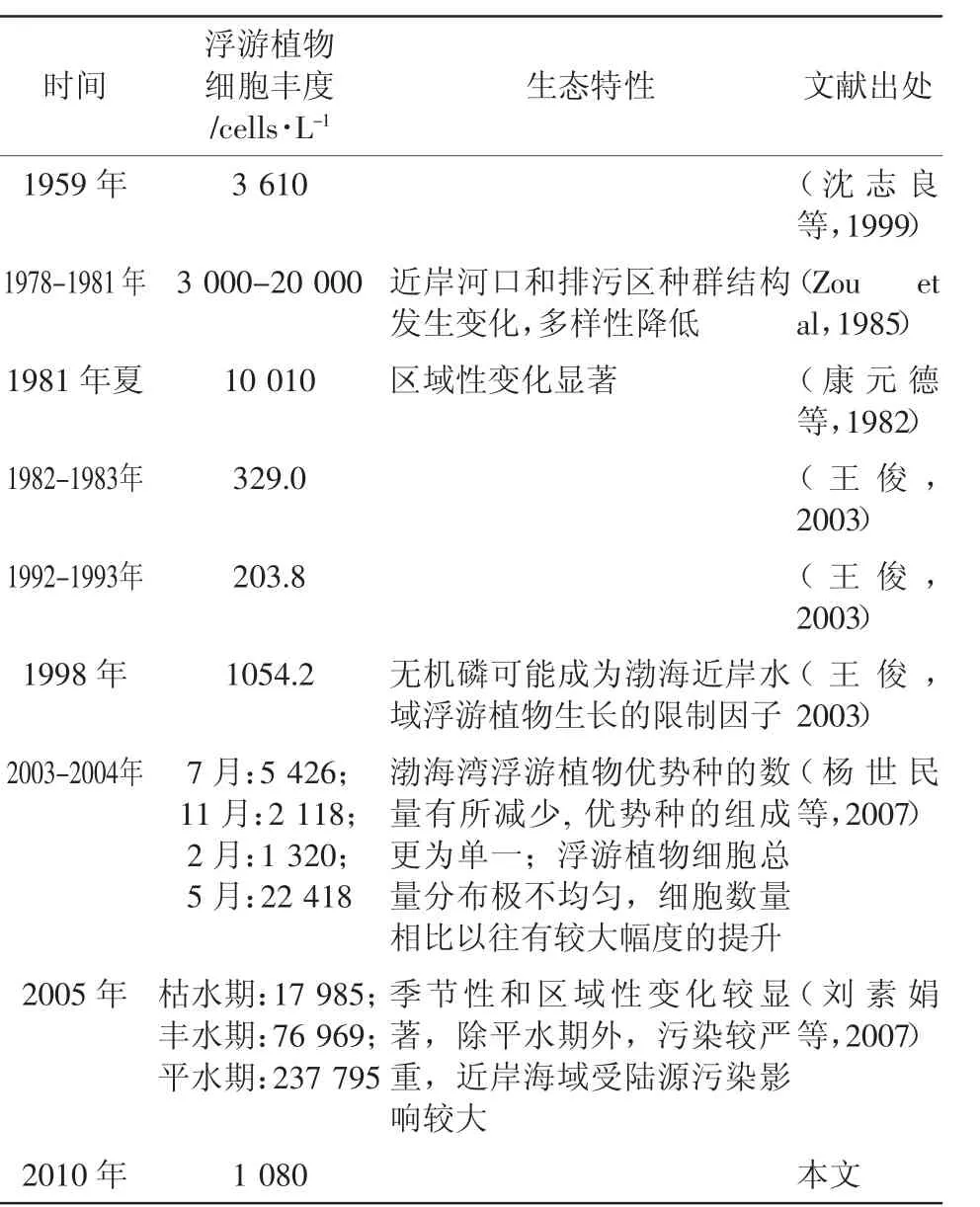
表6 渤海灣浮游植物生態特性的歷史變化
本文只是針對一個站位一次采樣的研究,可能存在一定的偶然性,實驗結果并不能完全代表海域的實際情況,但是在一定條件下揭示了浮游植物生長的重要規律,對于預測和控制海域富營養化具有重要意義。在進一步的研究中應加大采樣范圍和密度,同時采用更貼近現場環境的培養條件,并對浮游植物的種類變化加以細致的研究。
3 結論
2010年10月,對渤海灣西南部海域典型站位表層水體進行了營養鹽添加的模擬現場培養實驗,實驗判定了水樣的營養鹽限制類型,并在控制條件下,研究了不同氮磷比和不同硝酸鹽添加方式的生態響應,得到結果如下:
(1)單一添加磷酸鹽的實驗組浮游植物生長最明顯,且空白對照組磷酸鹽最早被耗盡,水樣體現了磷限制的特性。
(2)浮游植物的增長在總體上隨著氮磷比的降低而增大,最適宜氮磷比在5~15左右,略低于Redfield比。
(3)硝酸鹽的連續性添加比一次性添加更有利于浮游植物的生長,相對于高濃度沖擊性氮污染,低濃度持續性氮污染可能會產生更嚴重的生態影響,此處用一次性添加培養實驗可能會低估浮游植物的增長潛力。
了解海域的營養鹽限制特性對于海域水生態環境管理和修復具有重要的理論和實際意義,對于渤海灣的研究還需要大量和長期的實驗積累,本次實驗只是一個開始。但作為方法的創新,本文特色鮮明,在判定水樣營養鹽限制類型的基礎上,對培養實驗進行了改進,研究了水樣中浮游植物群落對于不同氮磷比和不同硝酸鹽添加方式的生態響應,并得到了較合理的結果。在進一步的研究中應加大采樣范圍和密度,同時采用更貼近現場環境的培養條件,并對浮游植物的種類變化加以細致的研究。
致謝感謝國家海洋局天津海洋環境監測站的石海明、徐玉山等同志在采樣期間的幫助。感謝天津大學和河北工程大學的徐曉甫、孫嬌、賈麗英、劉娟、劉巨波等同學在實驗期間的幫助與支持。
Dorth Q,Whitledge T E,1992.Does nitrogen or silicon limit phytoplankton production in the Mississippi river plume and nearby regions?Continental Shelf Research,12(11):1293-1309.
Fisher T R,Gustafson A B,Sellner K,et al,1999.Spatial and temporal variation of resource limitation in Chesapeake Bay.Marine Biology,133:763-778.
Hecky R E,Kilham P,1988.Nutrient limitation of phytoplankton in freshwater and marine environments:a review of recent evidence on the effects of enrichment.Limnology and Oceanography,33(4):796-822
Ho K C,Hodgkiss I J,1993.Assessing the limiting factors of red tide by bottle bioassay.Asian Marine Biology,10:77-94.
Hodgkiss I J,Ho K C,1997.Are changes in N:P ratios in coastal waters the key to increased red tide blooms?Hydrobiologia,352(1-3):141-147.
Justic D,Rabalais N N,Turner R E,et al,1995.Changes in nutrient structure of river-dominated coastal waters:stoichiometric nutrient balance and its consequences.Estuarine,Coastal and Shelf Science,40:339-356.
Nelson D M,Tréguer P,Brzezinski M A,et al,1995.Production and dissolution of biogenic silica in the ocean:Revised global estimates,comparison with regional data and relationship to biogenic sedimentation.Clobal Biogeochemical Cycles,9(3):359-372.
Tao J,2002.The eco-hydrodynamic environmental character of Bohai Bay.Proceedings of the Second international workshop on coastal eutrophication,China,1:15-24.
Zou J,Dong L,Qin B,1985.Preliminary studies on eutrophication and red tide problems in Bohai Bay.Hydrobiologia,127:27-30.
陳峰,姜悅,1999.微藻生物技術.北京:中國輕工業出版社.
鄧華健,陶建華,2004.浮游植物增長光響應動力學實驗研究.河北建筑科技學院學報,21(1):28-30,34.
胡俊,柳欣,張釩,等,2008.臺灣海峽浮游植物生長的營養鹽限制研究.臺灣海峽,27(4):452-458.
闞文靜,張秋豐,石海明,等,2010.近年來渤海灣營養鹽變化趨勢研究.海洋環境科學,29(2):238-241.
康元德,呂培頂,張坤誠,1982.渤海灣浮游植物光合作用強度的測定.海洋湖沼通報,4:47-51.
劉慧,董雙林,方建光,2002.全球海域營養鹽限制研究進展.海洋科學,26(8):47-53.
劉素娟,李清雪,陶建華,2007.渤海灣浮游植物的生態研究.環境科學與技術,30(11):4-10.
劉媛,曹振銳,黃邦欽,等,2004.東、黃海典型海區浮游植物對營養鹽添加的響應.廈門大學學報(自然科學版),43(增刊):147-152.
蒲新明,吳玉霖,2000.浮游植物的營養鹽限制研究進展.海洋科學,24(2):27-30.
沈志良,1999.渤海灣及其東部水域的水化學要素.海洋科學集刊,41:51-59.
孫霞,2005.光照對東海赤潮高發區赤潮藻類生長的影響.博士學位論文.青島:中國海洋大學.
唐松,2010.印度洋海水營養鹽添加模擬實驗中浮游植物生長的營養鹽限制作用.海洋科學,34(7):34-40.
王俊,2003.渤海近岸浮游植物種類組成及其數量變動的研究.海洋水產研究,24(4):44-50.
王曉偉,李純厚,李占東,等,2007.大亞灣海域春季浮游植物生長的限制性營養元素.中國水產科學,14(5):836-842.
王勇,趙澎,單寶田,2002.膠州灣營養鹽限制浮游植物生長的初步模擬現場實驗研究.海洋科學,26(10):55-59.
王勇,焦念志,1999.北黃海浮游植物營養鹽限制的初步研究.海洋與湖沼,30(5):512-518.
王玉玨,洪華生,王大志,等,2008.臺灣海峽上升流區浮游植物對營養鹽添加的響應.生態學報,28(3):1321-1327.
韋桂秋,2006.AGP試驗在珠江口監測的應用.碩士論文.青島:中國海洋大學.
蕭云樸,李揚,李歡,等,2009.溫州南麂列島海域硅藻、甲藻群落變化與環境因子的關系.海洋環境科學,28(2):167-169,201.
楊世民,董樹剛,李鋒,等,2007.渤海灣海域生態環境的研究Ⅰ.浮游植物種類組成和數量變化.海洋環境科學,26(5):442-445.
張均順,沈志良,1997.膠州灣營養鹽結構變化的研究.海洋與湖沼,28(5):529-535.
張平,沈志良,2001.營養鹽限制的水域性特征.海洋科學,25(6):16-19.
鄒立,張經,2001.渤海春季營養鹽限制的現場實驗.海洋與湖沼,32(6):672-678.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
