衢山島海域秋季浮游植物群落組成及影響因素
2012-08-14 05:48:48高瑜江志兵朱旭宇周青松陳全震曾江寧
海洋通報 2012年6期
高瑜,江志兵,朱旭宇,周青松,陳全震,曾江寧
(國家海洋局第二海洋研究所 國家海洋局海洋生態與生物地球化學重點實驗室,浙江 杭州 310012)
衢山島位于舟山群島的中北部,是我國第七大島,它處于長江、錢塘江入海口外緣,屬于舟山漁場的岱衢漁場海域。地理、水文、生物等優越自然條件,使岱衢漁場成為適宜多種魚類繁殖、生長、索餌、越冬的生活棲息地,其中以大黃魚尤為出名。但由于舟山群島位于長江下游,長江、錢塘江等陸地徑流帶入了大量的營養鹽,使得該海域富營養化十分嚴重(劉雪芹,2005),赤潮爆發頻繁。
浮游植物是海洋食物鏈中最基本的環節,是一些經濟魚類的重要餌料。國內對舟山漁場鄰近海域的浮游植物調查研究已有不少的報道。張賓等(2011)對舟山漁場氮磷營養鹽對浮游植物的限制進行了研究。魏娜等(2010)對舟山漁場浮游植物的時空分布以及優勢度進行了分析。劉子琳等(2001)分析了杭州灣—舟山漁場浮游植物的現存量并且計算了其初級生存力。朱根海等(2009)對衢山島西北部崎嶇列島的浮游植物和水環境狀況進行了研究。此外,還有不少學者對衢山島海域的漁業資源進行了研究(陳華等,2009;李建生等,2006)。但尚無專門針對衢山島海域浮游植物的研究,故筆者于2010年秋季大小潮期間對衢山島海域的浮游植物進行調查,并對調查結果做出分析,為深入了解該海區生態系統變化提供基礎數據。此外,舟山是浙江實行海洋經濟戰略的重點區域,舟山的海洋經濟必將迎來新的開發熱潮,其所面臨的海洋環境問題也必將更加突出,浮游植物的變化情況,可以作為水質監測以及評價的重要指標。因此,對該海域浮游植物的調查研究,具有重要的理論和實踐意義。
1 材料與方法
1.1 樣品的采集與處理
浮游植物樣品于2010年秋季大潮(10月22日-10月24日)和小潮(10月6日-10月14日)采集于衢山島鄰近海域23個大面站(圖1)。采用網目為76 μm的淺Ⅲ浮游生物網從底至表作垂直拖網采集浮游植物樣品,并用流量計計算濾水量。樣品用中性甲醛固定,經濃縮后用Leica DM2500顯微鏡觀察、鑒定和計數。水質分析指標分表底兩層采集,但考慮到表層指標與浮游生物有較大關系,實際分析時僅采用表層指標(陳雷等,2009)。水質分析項目有水溫、鹽度、可溶性磷酸鹽、可溶性無機氮、懸浮物等。水化學樣品采集、處理和測定以及浮游植物樣品的采集、貯存、運輸、預處理及實驗室分析測定均按《海洋監測規范》進行(國家質量技術監督局,1992,1999)。
1.2 數據處理
圖1 衢山島浮游植物采樣站位圖
分析浮游植物群落結構指數的計算方法(孫軍等,2003):(1)香農-威納多樣性指數(H′,Shannon-Wiener index)(2)物種優勢度,Y=ni·fi/N0;(3)Pielou均勻度指數,J=H′/log2S。式中:S為每站位所包含的總物種數;N為樣品中的總個體數;ni為樣品中第i種的個體數。
應用sufer 8.0繪制采樣站位圖以及細胞豐度圖。群落結構分析使用PRIMER 6.0軟件完成。
2 結果
2.1 浮游植物物種組成
大、小潮期間衢山島海域采共有浮游植物7門181種。其中,硅藻133種(占73.9%);甲藻38種(占21.1%),藍藻4種(2.2%),綠藻和金藻各2種(各占1.1%),黃藻和裸藻各1種(各占0.6%)。
大潮期間,共有浮游植物6門148種,其中硅藻109種(占73.6%),甲藻33種(占22.3%),藍藻和金藻各2種(各占1.4%),綠藻和裸藻各1種(各占0.7%);小潮期間,共有浮游植物7門114種,種類數低于大潮期間,其中硅藻80種(占70.2%),甲藻26種(占22.8%),藍藻3種(占2.6%),金藻2種(占1.8%),綠藻、黃藻和裸藻各1種(各占0.9%)。
2.2 浮游植物細胞豐度分布及優勢種
大潮期間,衢山島海域浮游植物細胞豐度為 2.21×104~19 637.25×104個/m3, 平 均 值 為2221.62×104個/m3。浮游植物細胞豐度高值區分布在大衢山北部黃澤山鄰近海域(D14、D15、D16、D18和D20站),該區細胞豐度平均值為9 800.82×104個/m3;浮游植物細胞豐度低值區位于大衢山至大長途島海域(S13、S14、S16、S17和S19站),平均值僅為17.69×104個/m3(圖2a)。
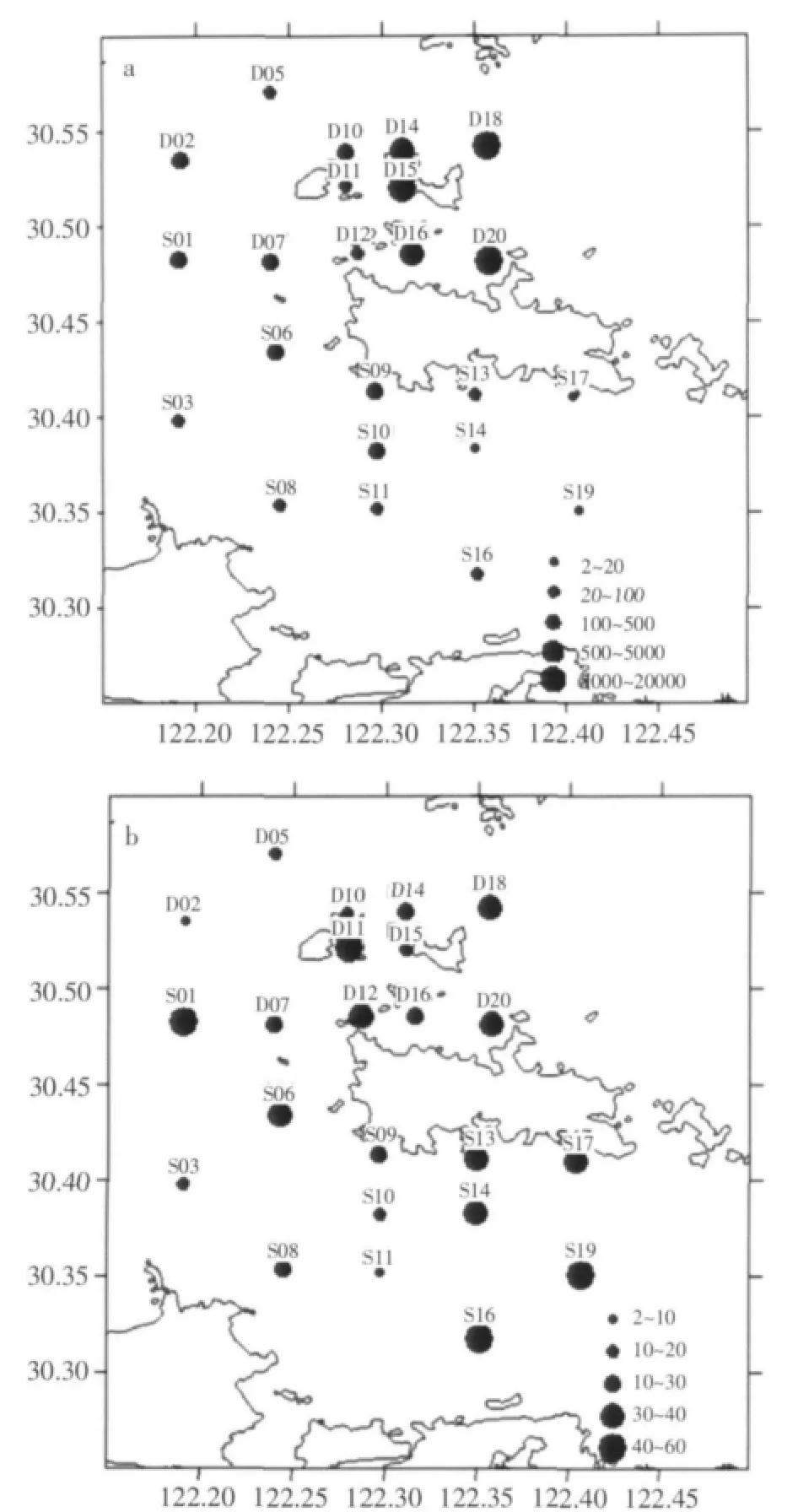
圖2 大小潮期間衢山島浮游植物細胞豐度(×104個/m3)(a.大潮;b.小潮)
小潮期間,衢山島海域浮游植物細胞豐度為5.90×104~58.10×104個/m3,平均值為 29.28×104個/m3。浮游植物細胞豐度分布高值區分布在大衢山至大長途島海域(S13、S14、S16、S17和S19站),平均值高達42.28×104個/m3;浮游植物細胞豐度低值區位于調查海域西北部(D02、D05、D10、D14和D15站位附近),平均值僅為17.14×104個/m3(圖2b)。
本次調查,衢山島秋季大潮期間浮游植物的絕對優勢種為中肋骨條藻(Y=0.987 2),這與魏娜等對舟山漁場海域的研究結果是相符合的。而本次調查區分了大小潮,小潮期間其優勢種為中肋骨條藻、瓊氏圓篩藻、整齊圓篩藻等。將大潮期間優勢種前兩位,小潮期間優勢種前五位列于表1。
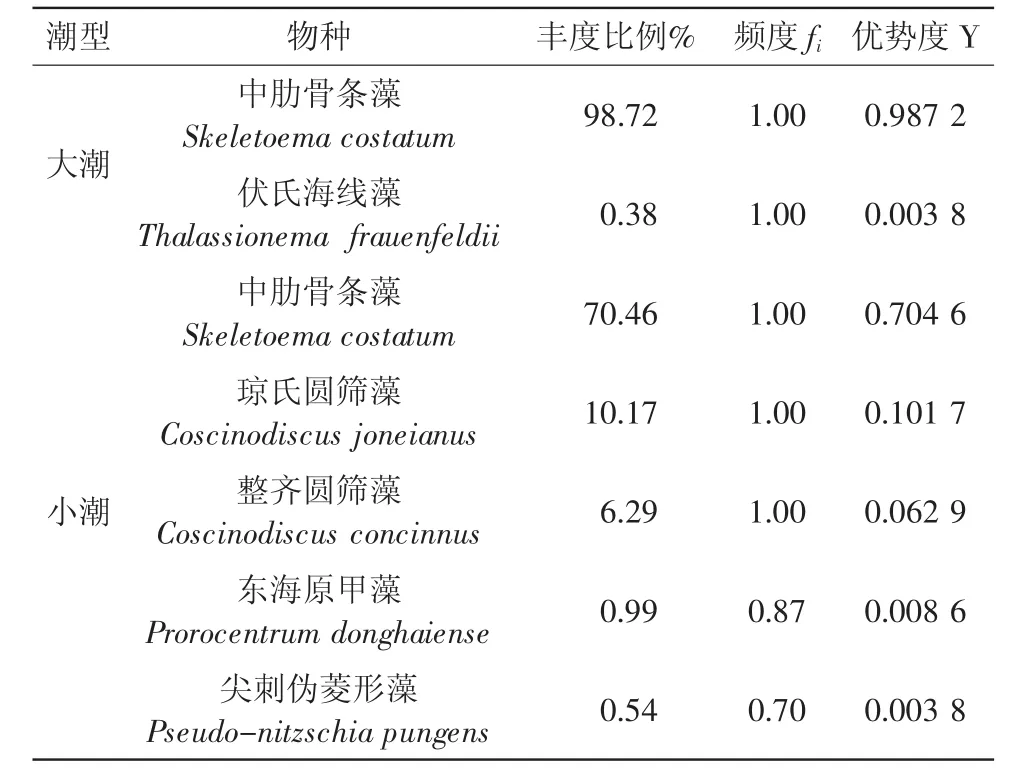
表1 調查站位浮游植物優勢種
本次調查海域浮游植物的數量分布和變化具有顯著的潮周期變化特點,表現在大潮期間浮游植物種類和細胞豐度明顯高于小潮期。大潮期間,細胞豐度的高值區集中于衢山島北部的D14、D15、D16、D18和D20站,而小潮期間,細胞豐度的高值區分布較為平均,且浮游植物的優勢種較為多樣。
2.3 群落多樣性分析
大潮期間調查區浮游植物的多樣性指數介于0.06~2.18,平均值為0.65,最低值出現在D20、D07、D16站;群落均勻度指數介于0.02~0.69,平均值為0.20。小潮期間此海域浮游植物的多樣性指數介于0.49~1.74,平均值為1.10,最低值出現在S16站;群落均勻度指數介于0.17~0.57,平均值為0.35。調查海域浮游植物的香農-威納多樣性指數、均勻度指數列于表2。
2.4 群落聚類分析
分別對大小潮23個站位的浮游植物進行Cluster聚類(圖3、圖4)。Cluster結果以40%的群落結構相似性來劃分,則23個站位大潮期間可分為5組,其中D14、D16、D20、D18、D15分為一組,S19和S14 各為一組,S17、S11、S16、D11、S13、S08 分為一組,其余站位分為一組;小潮期間可分為4組,其中S17和D10各為一組,D02、S03、S14、S06、S11、D14、D12分為一組,其余站位分為一組。
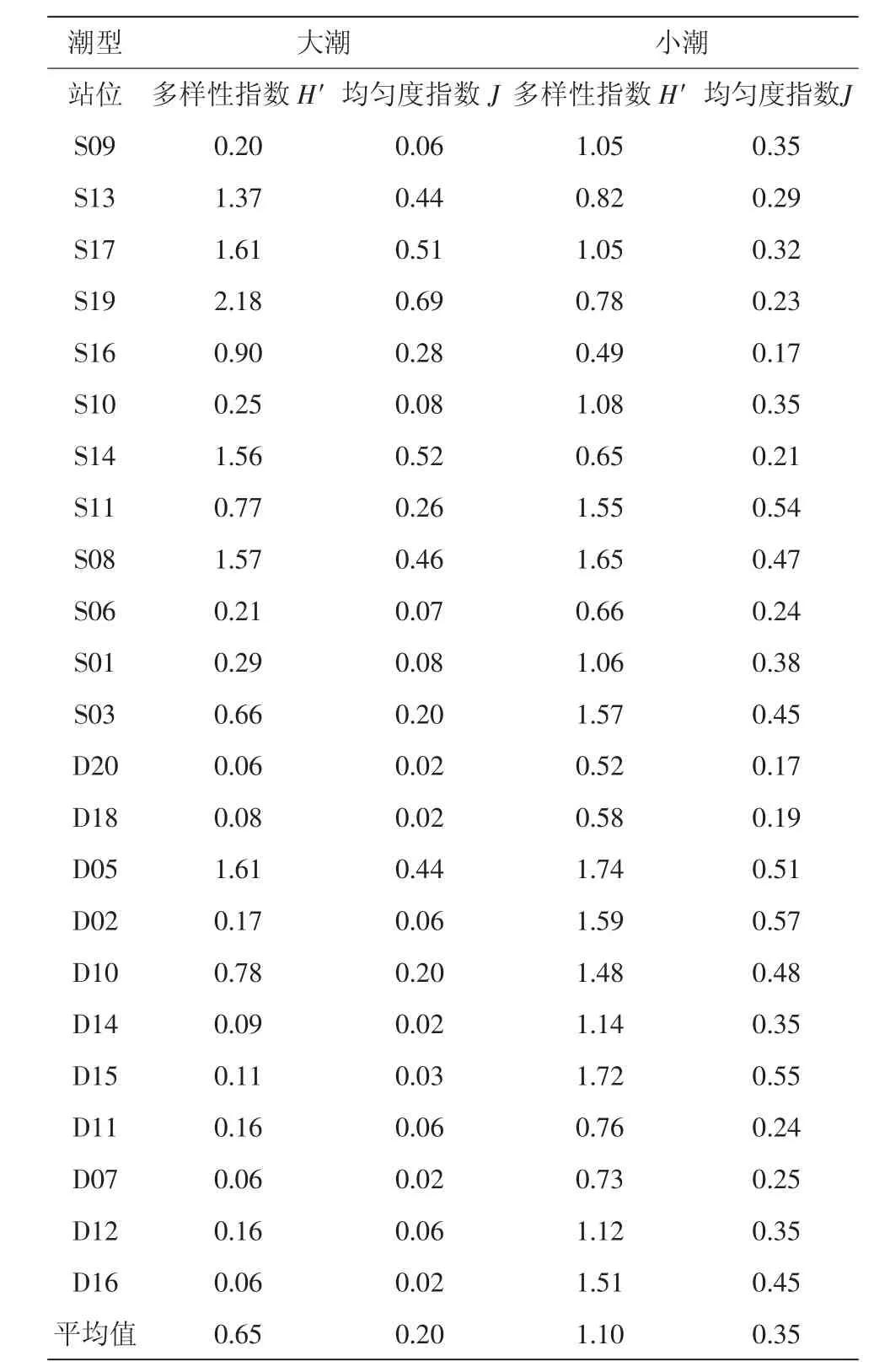
表2 大小潮期間衢山島海域浮游植物群落生物多樣性指數
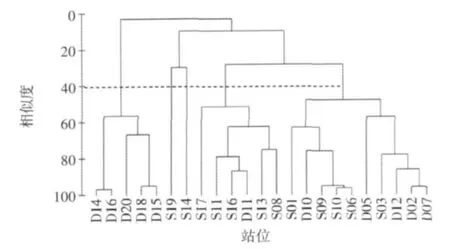
圖3 大潮期間衢山島海域浮游植物群落的聚類圖
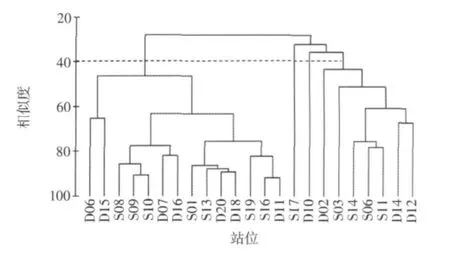
圖4 小潮期間衢山島海域浮游植物群落的聚類圖
大潮期間,群落相似性呈明顯斑塊分布,D14、D16、D20、D18、D15 5個站位是浮游植物細胞豐度的高值區,位于衢山島的北部,S19和S14雖在聚類中分屬不同的類群,但都是浮游植物細胞豐度的低值區,且位于衢山島的東南部;小潮期間,群落相似性分布的規律性不明顯。由此可見,潮汐對浮游植物的種類組成和細胞豐度都有著顯著影響。
2.5 衢山島海域水質情況
大潮期間,衢山島海域無機氮分布情況為衢山島北部高于南部(圖5a),平均濃度為0.81 mg/L,最高值位于D16站位,達1.11 mg/L;無機磷含量分布較為均勻(圖5b),平均濃度為0.04 mg/L。
小潮期間,無機氮分布同樣為衢山島北部海域稍高于南部(圖5c),平均濃度為1.04 mg/L,最高值位于D10站位,達1.18 mg/L;無機磷含量與大潮期無較大差異(圖5d),平均值為0.04 mg/L。
根據中華人民共和國海水水質標準(GB 3097-1997),衢山島海域無機氮含量遠遠超過四類水標準(無機氮<0.50 mg/L),活性磷酸鹽含量也僅僅是三類水范圍。由此可見,衢山島海域富營養化程度較為嚴重。
在本次調查中發現,大小潮對衢山海域的溫度、鹽度、pH影響極小,無機氮和活性磷酸鹽的含量在大小潮期間的差異也并不顯著,但是懸浮物濃度差異卻極為明顯。大潮期間,衢山島海域懸浮物濃度平均值為464.02 mg/L,而小潮期懸浮物平均濃度僅為35.16 mg/L。故僅選取無機氮、活性磷酸鹽及懸浮物3個參數與浮游植物豐度做相關性分析。結果表明(表3),衢山島海域浮游植物豐度與無機氮、無機磷含量無顯著相關關系,但與懸浮物濃度呈顯著的負相關關系。
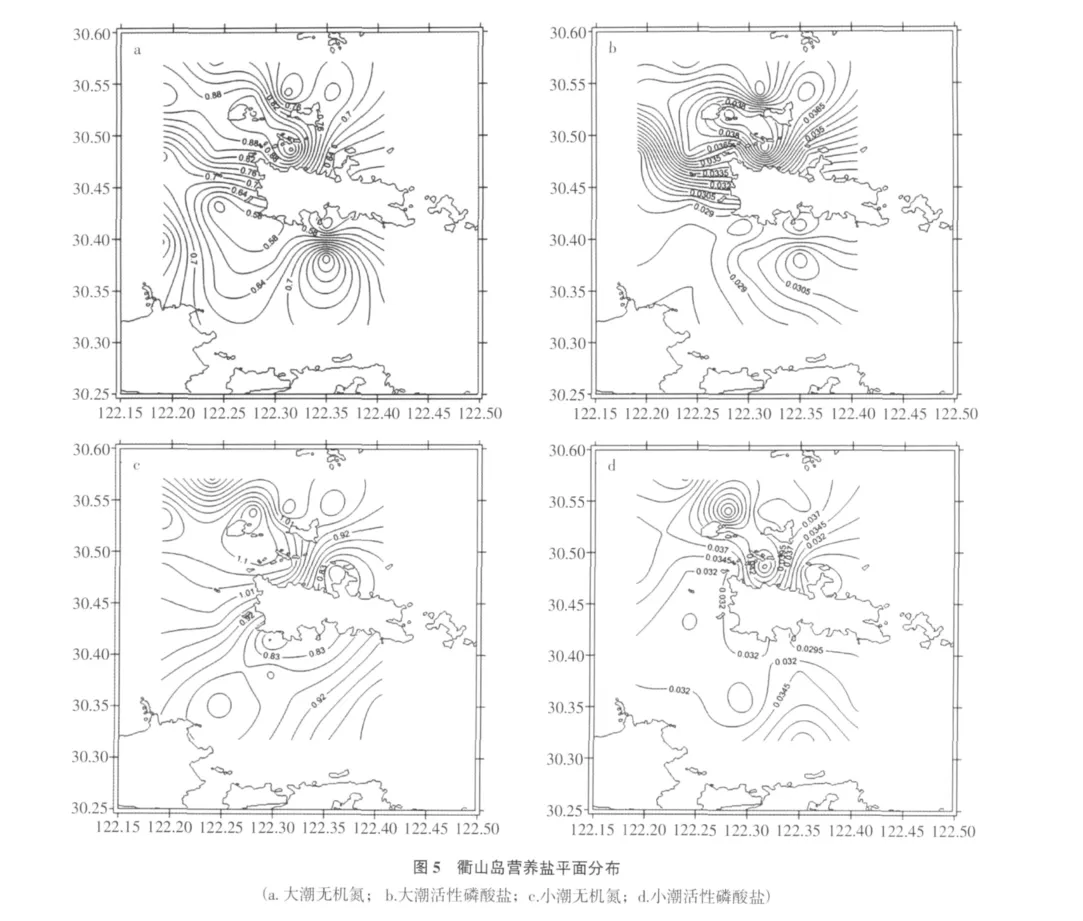
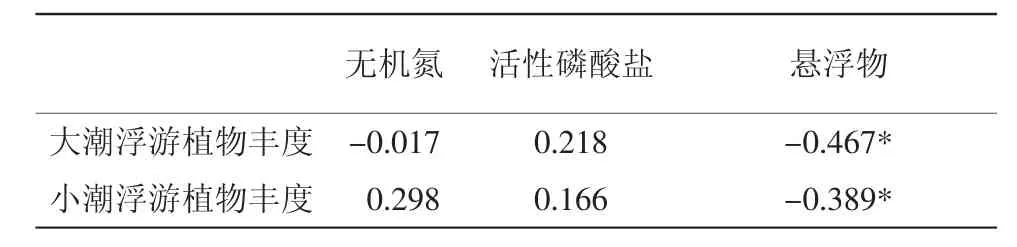
表3 大小潮期間浮游植物豐度與環境因子相關系數
3 討論
3.1 浮游植物群落與水團、營養鹽等環境因子的關系
衢山島附近海域至少存在3個主要水團,分別為江浙沿岸水、臺灣暖流表層水和黃海混合水(張啟龍等,2007)。江浙沿岸水低溫低鹽,而臺灣暖流則是高溫高鹽,寒暖流在此交匯,使得此區域溫度適宜,非常適合浮游植物的生長,從而為漁場的形成創造了良好條件。
本海域營養鹽的來源一方面是由于長江徑流攜帶大量陸源營養物質,另一方面臺灣暖流的入侵也為本海域提供了磷酸鹽的補充(Chung et al,1999),因而營養鹽并不是該海域浮游植物生長的限制因子。大潮期間,調查海域水體渾濁,透明度低,水環境條件極不穩定,多數種類很難繁殖生長,但是中肋骨條藻卻占據了細胞豐度98%的比例,說明中肋骨條藻適宜偏淡水低鹽環境,尤其是在徑流量偏大水域(唐峰華等,2010)。
根據文獻資料(徐兆禮 等,1999;楊世民,2006;王全喜等,2008)及水團因素,將調查區浮游植物分為:(1)近岸種,代表種為中肋骨條藻Skeletoema costatum和瓊氏圓篩藻Coscinodiscus joneianus以及伏氏海線藻 Thalassionema frauenfeldii等,在此次調查出現的浮游植物中,這些種類是數量上具有舉足輕重的優勢種;(2)淡水種,代表性種為黃絲藻Triboneema sp.、顫藻Oscillatoria sp.、具孔盤星藻Pediastrum clathratum、單角盤星藻Pediastrum simplex等。衢山島海域位于長江沖淡水的稀釋區域,這些淡水種主要由長江徑流輸入,但秋季為長江平水期,故本類浮游植物出現數量均不大;(3)外海高鹽種,代表種為虹彩圓篩藻Coscinodiscus oculus-iridis、筆尖形根管藻Rhizosolenia styliformis、密連角毛藻Chaetoceros densus等,本類群則由外海水攜帶而來。
3.2 浮游植物多樣性分析
浮游植物群落的香農-威納多樣性指數、均勻度指數可以作為評價環境質量的依據(陳碧鵑等,2001)。物種組成的多少及每個物種個體數量是衡量生物群落多樣性的基礎,且各個物種間的均勻程度也是衡量多樣性的重要因素(Magurran,1988)。《水生生物監測手冊》(國家環保局水生生物監測手冊編委會,1993)中指出,當0<H′<1時,表明水體受到人為的重度影響;當H′=1~3時,表明水體受到人為的中度影響。此外,實踐應用中,當均勻度J>0.3時,表明海區內浮游植物的多樣性較好(馬建新等,2002)。現取H′<1且J<0.3作為多樣性較差的標準對各站位進行綜合評價:大潮期間,除 S13、S17、S19、S14、S08、D05 6 個站外,其他站位的多樣性均較低;小潮期間,群落的多樣性較大潮有所好轉,只有S13、S19、S16、S14、S06、D20、D18、D11、D07站位多樣性水平較低。當優勢種較多,或無優勢性特別突出的種類,此時浮游植物群落結構是穩定的(Pinckney et al,1998),所以,小潮期衢山島海域浮游植物群落穩定性優于大潮期。
3.3 潮周期對浮游植物群落的影響
衢山島海域浮游植物的種類和細胞豐度具有明顯的潮周期變化特征,大潮期間浮游植物種類以及細胞豐度均大于小潮期,且大潮期間浮游植物總量斑塊分布現象比小潮期間明顯。衢山島所處海域位于以強潮和流作用為主要特點的強動力環境中,漲、落潮歷時受到長江和錢塘江入海徑流的顯著影響(陳沈良,2000),水動力實際上對本海區浮游植物的分布起著一種輸移和聚集的作用(顧新根等,1995)。大潮期潮流急、流量大,在此潮流作用下位于外海浮游植物向近岸擴展,而小潮期間潮流流速相對較緩,此時浮游植物隨水流向外海擴散,由此導致了浮游植物在大小潮期間數量的差異。
4 結論
(1)大、小潮期間衢山島海域共鑒定出浮游植物7門180種。其中,硅藻133種,占總種數的73.89%,是衢山島海域浮游植物群落的主要類群;甲藻38種,占總種數的28.50%;藍藻4種,綠藻和金藻各2種,黃藻和裸藻各1種。大潮期浮游植物種類數大于小潮期。
(2)大潮期間,衢山島海域浮游植物平均細胞豐度為2 221.62×104個/m3,中肋骨條藻為絕對優勢種;小潮期間,浮游植物平均細胞豐度為29.28×104個/m3,此時出現了中肋骨條藻、瓊氏圓篩藻、整齊圓篩藻、東海原甲藻等較為多樣的優勢種。
(3)大潮期間,衢山島海域的多樣性指數、均勻度指數、豐富度指數的均值分別為0.65、0.20、2.56;而小潮期間,其三大指數的均值分別為1.10、0.35、2.75。因此總體來說,衢山島海域小潮期間的生物多樣性和豐富度尚好,群落結構較為穩定。但大潮期間的生物多樣性較差。
(4)通過聚類分析,大潮期間衢山島海域浮游植物群落可以分為5個類群。小潮期間可以分為4個類群。不同類群之間差異性顯著。
(5)影響衢山島海域浮游植物種類和數量分布的決定性因素是潮汐,光照是影響浮游植物豐度的重要因素,而營養鹽并沒有成為該海域浮游植物生長的限制因子。
Chung S W,Jan S,Liu K K,2001.Nutrients fluxes through the Taiwan Strait in spring and summer 1999.J Oceanogr,57(1):47-53.
Magurran A E,1988.Ecological diversity and its measurement.Sydney:Croom Helm,54:7-46.
Pinckney J L,Paerl H W,Harrington M B,et al,1998.Annual cycles of phytoplankton community-structure and bloom dynamics in the Neuse River Estuary,North Carolina.Marine Biology,131(2):371-381.
陳碧鵑,陳聚法,崔毅,等,2001.萊州灣東部養殖區浮游植物的生態特征.海洋水產學研究,22(3):64-70.
陳華,徐兆禮,2009.舟山漁場衢山島海域春夏季魚類數量變化.海洋漁業,31(2):179-185.
陳雷,徐兆禮,姚煒民,等,2009.甌江口春季營養鹽、浮游植物和浮游動物的分布.生態學報,29(3):1571-1577.
陳沈良,2000.崎嶇列島海區的水文泥沙及其峽道效應.海洋學報,3(22):123-131.
顧新根,袁騏,沈煥庭,等,1995.長江口最大渾濁帶浮游植物的生態研究.中國水產科學,2(1):16-27.
國家環保局水生生物監測手冊編委會,1993.水生生物監測手冊.南京:東南大學出版社.
國家質量技術監督局,1999.海洋監測規范第4部分:海水分析(GB17378-1998).北京:中國標準出版社.
國家質量技術監督局,1992.海洋監測規范第6部分:海洋生物調查(GB/T 12763-1991).北京:中國標準出版社.
李建生,李圣法,程家驊,2006.長江口漁場魚類組成和多樣性.海洋漁業,28(1):37-41.
劉雪芹,2005.舟山近岸海域富營養化評價.海洋湖沼通報,2:55-60.
劉子琳,寧修仁,蔡昱明,2001.杭州灣—舟山漁場秋季浮游植物現存量和初級生產力.海洋學報,23(2):93-99.
馬建新,鄭振虎,李云平,等,2002.萊州灣浮游植物分布特征.海洋湖沼通報,4:63-67.
孫軍,劉東艷,寧修仁,等.2003.2001/2002年夏季南極普里茲灣及其鄰近海域的浮游植物.海洋與湖沼,34(5):519-532.
孫軍,劉東艷,2002.中國海區常見浮游植物種名更改初步意見.海洋與湖沼,33(3):271-286.
唐峰華,伍玉梅,樊偉,等,2010.長江口浮游植物分布情況及與徑流關系的初步探討.生態環境學報,19(12):2934-2940.
王全喜,曹建國,劉妍,等,2008.上海九段沙濕地自然保護區及其附近水域藻類圖集.北京:科學出版社.
魏娜,胡顥炎,毛宏躍,等,2010.舟山漁場及其鄰近海域浮游植物生態調查與研究.海洋環境科學,29(2):170-173.
徐兆禮,白雪梅,袁騏,等,1999.長江口浮游植物生態研究.中國水產科學,5(6):52-54.
楊世民,董樹剛,2006.中國海域常見浮游硅藻圖譜.青島:中國海洋大學出版社.
張賓,章飛軍,郭遠明,等,2011.舟山漁場海域春夏季氮磷分布及其對浮游植物的限制.浙江海洋學院學報(自然科學版),30(3):185-189.
張啟龍,王凡,趙衛紅,等,2007.舟山漁場及其鄰近海域水團的季節特征.海洋學報,29(5):1-9.
朱根海,施青松,張健,等,2009.崎嶇列島附近海域浮游植物與水環境狀況研究.海洋學報,31(4):149-158.
