Follistatin、Activin A與BMP-4在大鼠腦發育過程中的表達及意義
2012-07-31 09:23:00盧彥春穆長征李偉偉
山東醫藥 2012年35期
關鍵詞:海馬
盧彥春,張 蕾,穆長征,李偉偉,金 輝
(1遼寧醫學院生物化學教研室,遼寧錦州 121000;2遼寧醫學院附屬第三醫院)
研究表明,多種細胞因子及信號通路共同參與神經分化的調控機制,其中卵泡抑素(FS)能夠誘導神經分化,并拮抗激活素(Activin)的生物活性進一步抑制胞內的Smad3的信號激活[1]。有學者認為,FS作為Activin特異結合蛋白,還可與TGF-β家族的其他成員結合,如骨形態發生蛋白(BMPs)、myostatin等,并抑制BMP信號而達到神經誘導作用的。而最新研究表明,FS沒有阻止BMP信號,其功能主要是作為 Activin抑制劑,而不是 BMP配體[2]。2010年4月~2012年3月,我們通過觀察鼠神經發育不同階段和部位FS、Activin A與BMP-4基因的表達變化及定位情況,以探討神經細胞誘導和分化的可能機制。
1 材料與方法
1.1 動物及分組 取同期受孕SD大鼠30只,由遼寧醫學院科學實驗動物中心提供。將30只大鼠按照胎鼠發育時間分為胚胎8.5 d(E8.5組)、13 d(E13組)、18 d(E18組)及出生后3 d(P3組)、7 d(P7組)、30 d(P30組)各5只。
1.2 FS、Activin A、BMP-4 表達檢測 E8.5 組、E13組、E18組中每只孕鼠取5只胎鼠,斷頭取腦,經40 g/L(4%)多聚甲醛固定;P3組、P7組、P30組各取5只幼鼠經腹腔麻醉,打開胸腔,用300 mL(0.1 mol/L)PBS(pH7.4)經左心室灌注沖洗,再用40 g/L(4%)多聚甲醛磷酸鹽緩沖液300 mL緩慢灌注固定。應用腦立體定位儀分別切取包含皮質、紋狀體、海馬、嗅球組織的腦片,石蠟包埋,連續進行冠狀切片(厚度為5 μm),每4張取1張,行免疫組化染色。觀察 FS、Activin A、BMP-4 在皮質、紋狀體、海馬、嗅球處表達。免疫組化染色操作步驟按SABC免疫組化染色試劑盒操作說明進行。
1.3 統計學方法 所得數據輸入計算機,用SPSS for Windows 10.0版本軟件進行t檢驗和方差分析,結果以±s表示。檢驗水準α=0.05。
2 結果
2.1 皮質 各組皮質陽性細胞分布均勻,FS與Activin A在E8.5組陽性細胞數最多,在E13組皮質的錐體細胞層,細胞著色較深,提示此層細胞FS(圖1)與Activin A(圖2)含量較多,P7組與P30組陽性細胞較少且著色最弱。BMP-4在E8.5~P7組陽性細胞較少,在P30組陽性細胞顯著增多,可見胞質染色較強的細胞,主要分布于顆粒層和錐體細胞層(圖3)。
2.2 紋狀體 FS與Activin A在E8.5組陽性細胞數最多,E13組與E18組陽性細胞數減少且著色較強,可見胞質染色較強的細胞,主要分布于顆粒層(FS見圖4、Activin A見圖5),P3組陽性細胞數低于E18組,P7組和P30組陽性細胞較少且著色最弱。BMP-4在E8.5組到P7組陽性細胞較少,著色相對較弱,在P30組陽性細胞顯著增高,細胞著色深,主要分布于顆粒層(圖6)。
2.3 海馬 FS與Activin A在E8.5組陽性細胞數最多,E13組明顯少于E18組,齒狀回陽性細胞體積較小,數目較多,E8.5組到P7組各亞區陽性細胞數都較多,細胞著色深,主要分布在錐體細胞層、顆粒層(FS見圖7、Activin A見圖8),P30組陽性細胞數進一步減少。BMP-4在E8.5組到P7組陽性細胞數極少且著色極弱,P30組陽性細胞著色較強,細胞著色深,集中在錐體細胞層、顆粒層(圖9)。
2.4 嗅球 FS和Activin A在E8.5組陽性細胞最多。E13組陽性細胞數略減少,E18組與P3組陽性細胞數略少于E13組,分布于各層,胞質呈深棕黃色,核空泡狀,僧帽細胞可見陽性突起(FS見圖10、Activin A見圖11),P7組到 P30組進一步減少。BMP-4在E8.5組到P7組陽性細胞數較少,P30組陽性細胞增多,呈高表達,陽性細胞均在纖維層、僧帽細胞層、顆粒細胞層、髓層較多(圖12)。
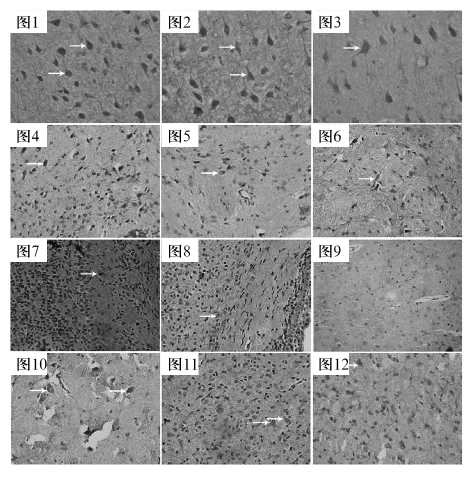
圖1 ~12 大鼠大腦皮質、紋狀體、海馬、嗅球內FS、Activin A與BMP-4免疫組化染色(×400)
3 討論
研究表明,多種細胞因子及信號通路共同參與神經分化的調控機制,成體神經發生亦受缺血、運動等眾多因素的影響[3,4]。其中,Fs能夠誘導神經分化,并拮抗Activin的生物活性進一步抑制胞內的Smad3 的信號激活[1]。Shoji-Kasai等[5]發現,體外培養的海馬神經元突觸數量和樹突長度的增加有賴于Activin A。新近的實驗表明,Activin A具有促進原代培養背根神經節神經元神經突起生長、抑制神經損傷分子NO釋放作用,與神經生長因子具有協同效應[6,7]。根據配體中的半胱氨酸殘基的分布將TGF-β家族成員區別為三個亞群,即標準TGF-βs家族、activin/inhibins家族、BMPs家族。Fs與 Activin和Myostatin以高親和力結合形成基本不可逆的復合物,阻止了受體配體的相互作用,在脊椎動物中幾個作為TGF-β配體亞家族的拮抗劑均與Fs有關。
Bickel等發現,BMP和Activin配體均為FS作用的靶點。而最新研究表明,FS沒有阻止BMP信號,其功能主要是作為Activin抑制劑,而不是BMP配體,這與Bickel的研究不同。本實驗發現大鼠大腦皮質和紋狀體內FS、Activin A在E8.5組陽性細胞數最多,在E13組與E18組陽性細胞數減少,P3組陽性細胞數低于E18組,P7組及P30組細胞著色最弱,而BMP-4基因呈現相反的表達,在E8.5組至P3組著色相對較弱,在P7組開始增強,在P30組陽性細胞顯著增高,提示Fs對 ACT的拮抗特點與Bickel研究結果相吻合,而Fs與BMP-4的表達呈相反波動趨勢。
眾所周知,神經板是最早出現的神經系統發育的結構。參與神經板發生的主要有BMP、Fs、Chorclin和Noggin。Fs是一類分泌性因子,表達于原腸胚期的組織原區及動物極性外植體中。新近研究發現,皮層、紋狀體、黑質、杏仁核、海馬CAI區、腦干背側迷走神經復合體、室旁核都有神經發生[8]。本文結果表明,FS、Activin A在大鼠海馬E8.5組陽性細胞數最多,E13組明顯低于E8.5組,而BMP-4在此時的著色極弱,提示FS與Activin A蛋白能夠抑制BMP-4的作用,可能對胚胎期和成年期海馬的神經發生調控起重要作用。近來研究也表明,在出生后20 d至成年階段齒回能產生顆粒細胞,然而和胚胎期及新生期相比,數量明顯減少[8]。成年大鼠神經元前體細胞的遷移流主要有兩條路徑:一條是從SVZ到嗅球的RMS;另一條是位于皮層和胼胝體周圍遷移流[3]。研究發現,大鼠嗅球存在兩次嗅感覺神經細胞凋亡高峰,分別在胚胎13 d和出生后第5天,隨著嗅球的神經發生,嗅感覺神經細胞發育成熟[9,10]。本研究發現,FS 和 Activin A 在 E8.5 組嗅球陽性細胞分布于各層,著色最強。E13組陽性細胞數略減少,而BMP-4在此時陽性細胞數較少。FS和ACT在E18組至生后P3組呈中等強度表達,陽性細胞數略少于E13組,二者在P7組進一步減少,P30組降至最低。而BMP-4在P7組陽性細胞數較少,P30組陽性細胞增多,呈高表達,由此推測FS和Activin A蛋白能夠抑制BMP-4的作用從而阻止嗅上皮的嗅感覺神經細胞凋亡的發生。
總之,FS在神經誘導的調控機制的過程中與Activin A和BMP-4密切相關,作為外胚層的內部因素在正常發育中有重要意義。但神經發育調控機制復雜,尚需進一步的探索研究。
[1]李偉偉,張蕾,田力.卵泡抑素誘導骨髓間充質干細胞分化為神經元樣細胞[J].神經解剖學雜志,2009,25(6):616-620.
[2]Pentek J,Parker L,Wu A,et al.Follistatin preferentially antagonizes activin rather than BMP signaling in drosophila[J].Genesis,2009,47(4):261-273.
[3]張濤,王景周.成體腦內神經發生和腦缺血對其影響[J].中華神經醫學雜志,2009,8(2):207-210.
[4]王紅星,徐冬晨,王彤.中樞神經再生策略:增強神經因子介導的再生環境“允許”作用[J].中國康復醫學雜志,2008,23(4):382-384.
[5]Shoji-Kasai Y,Ageta H,Hasegawa Y,et al.Activin increases the number of synaptic contacts and the length of dendritic spine necks by modulating spinal actin dynamics[J].Cell Sci,2007,120(Pt 21):3830-3837.
[6]方琳,柳忠輝,劉永茂.激活素促進雞胚神經節神經突起生長作用[J].動物學雜志,2006,41(6):23-28.
[7]方琳,劉永茂,葛敬巖.激活素結合蛋白阻斷激活素誘導雞胚神經節神經突起生長作用及其機制研究[J].中風與神經疾病雜志,2007,24(4):395-397.
[8]Raymond AD,Kucherepa NN,Fisher KR,et al.Neurogenesis of oxytocin-containing neurons in the paraventricular nucleus(PVN)of the female pig in 3 reproductive states:puberty gilts and lactating sows[J].Brain Res,2006,1102(1):44-51.
[9]West MJ,Slomianka L,Gundersen HL.Unbiased stereological estimation of the total number of neurons in the subdivision of the rat hippocampus using the optical fractionator[J].Anat Rec,1991,231(4):482-497.
[10]Cowan CM,Roskams AJ.Caspase 3 and Caspase 9 mediate developmental apoptosis in the mouse olfactory system[J].J Comp Neurol,2004,474(1):136-148.
猜你喜歡
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
娃娃樂園·綜合智能(2020年8期)2020-08-28 00:32:14
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
作文周刊·小學二年級版(2018年9期)2018-04-18 10:01:40
小學生導刊(2018年1期)2018-03-15 08:02:37
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
科技知識動漫(2016年6期)2016-06-24 21:04:53
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
汽車觀察(2009年1期)2009-02-18 09:11:50
