6種野生鳶尾種皮、種苗特征與進化的關系1)
2012-06-13 06:20:50陳士惠卓麗環
東北林業大學學報 2012年1期
孫 穎 陳士惠 卓麗環 王 玲
(東北林業大學,哈爾濱,150040) (上海農林職業技術學院) (東北林業大學)
鳶尾屬是單子葉植物鳶尾科中最大的屬,也是最進化的屬。野生的鳶尾屬植物生命力強,抗逆性強,綠色期長,觀賞價值高,有大面積推廣應用的前景。目前世界鳶尾屬的研究主要集中在雜交育種、創制新品種上[1-5]。有關系統發育理論方面的研究,在國外的鳶尾屬植物相關文章中有一定涉及[6-8],國內研究的較少[9-10]。對于我國東北野生鳶尾的種子相關結構的研究,將為更廣闊地區乃至全國的鳶尾資源的系統研究和生產應用提供部分理論依據。
1 試驗材料
試驗材料于東北林業大學帽兒山實驗林場、哈爾濱師范大學家屬區院內、沈陽市植物園的鳶尾園采集(表1)。根據文獻[11]所做的對60種鳶尾的數量分類學研究,將中國鳶尾屬植物劃分為11類,從中可以看出,本研究所選出的東北鳶尾均屬于第一類無附屬物亞屬12個種中的6種。這是鳶尾屬中最原始的類群,彼此間的親緣關系較近,是一個很自然的類群。
2 試驗方法
2.1 種皮結構觀察
采用日本大學JEOL公司生產的JSM—5610LV型掃描電子顯微鏡進行觀察,每個種觀察3粒,并選擇圖像清晰、具代表性區域的畫面拍照。拍照部位均為種子背面,種皮雕紋紋飾分別放大300倍及1 000倍。

表1 材料來源
2.2 種苗發育過程觀測
采用常規發芽試驗。出苗期觀察萌發的種子,每種30粒,記錄其發育過程,記錄胚根、子葉鞘、真葉出現的時間。于萌發后7 d測量胚根、子葉鞘、子葉聯結的長度。
3 結果與分析
3.1 種皮超顯微結構
觀察結果表明,6種鳶尾種皮超顯微形態特征差異明顯。玉蟬花初級雕紋為條網狀紋飾。網眼凹陷較深,由長條狀的四邊形或五邊形組成。網脊凸起,較粗,高度一致。各網眼的網脊界限清晰。次雕紋由次生壁加厚的不規則波狀乳突構成(圖1(a)、圖1(b))。溪蓀初級雕紋為條網狀紋飾。網眼較平整,由不規則的長條狀五邊形或六邊形組成,近似魚鱗狀。網脊為單條紋,狹窄,隆起程度差。各網眼的網脊界限清晰。次雕紋較光滑(圖1(c)、圖1(d))。北陵鳶尾初級雕紋為網狀紋飾。網眼凹陷,由近規則的正五邊形和正六邊形組成。網脊突出,但沒有玉蟬花的網脊明顯。網脊較粗,高度不一致。各網眼的網脊界限較清晰,個別地方模糊。次雕紋由次生壁加厚的不規則條狀網紋構成(圖1(i)、圖1(j))。馬藺初級雕紋為蜂窩狀紋飾。網眼凹陷,由較為規則的正五邊形或六邊形組成。網脊凸出,較粗,寬度一致,高度近一致。各網眼的網脊界限十分清晰。次雕紋較光滑,個別為由次生壁加厚的不規則疣狀乳突構成(圖1(k)、圖1(l))。黃菖蒲初級雕紋為條網狀紋飾。網眼凹陷,由長條狀的四邊形或五邊形組成。網脊凸起,較粗,高度一致。各網眼的網脊界限清晰。次雕紋由次生壁加厚的均勻分布的疣狀乳突構成(圖1(e)、圖1(f))。燕子花初級雕紋為條網狀紋飾。網眼較平整,由不規則的長條狀五邊形或六邊形組成,近似魚鱗狀。網脊較窄,隆起程度差,高度一致。各網眼的網脊界限清晰。次雕紋由次生壁加厚的不規則條狀網紋構成(圖1(g)、圖1(h))。
3.2 出苗期種苗結構特征
種子萌發時,下胚軸不發育或不伸長,只有上胚軸和胚芽迅速向上生長,形成幼苗的主莖,而子葉始終留在土壤中,這種方式形成的幼苗,稱為子葉留土幼苗。本文所研究的6種鳶尾科植物均為子葉留土型。這種類型植物的子葉明顯區別出子葉鞘、子葉聯結和吸器。子葉鞘起著保護胚芽的作用,吸器是吸收胚乳的營養物質,子葉聯結起著前兩者之間聯系的作用。更進化的種類在此基礎上發育出另一些鞘狀結構,稱為舌狀鞘、假舌狀鞘。
解剖鏡下觀測發現,鳶尾屬植物種子萌發時胚突破種皮的一端為圓鈍形,當露白達到2~3 mm時,胚逐漸變尖,呈錐狀,這是形成的胚根。這種現象與種胚在種子內部的狀態有關。在種子內,種胚靠近發芽口的一側即為圓鈍狀,遠離發芽口的一側呈錐狀。大部分玉蟬花、黃菖蒲的胚根密生根毛,溪蓀和北陵的根部具有較密根毛的比例很低,只有28%的馬藺根部生有根毛。在對鳶尾科射干屬射干的同類研究中發現,旱生的射干萌發后,只有16%的胚根生有較顯著的根毛。由此認為:旱生的鳶尾趨向于根毛稀疏,而濕生種類則趨向于密生根毛。
觀察種苗可以看出(表2):溪蓀、北陵、黃菖蒲、燕子花子葉只有基鞘,沒有舌狀鞘,這4種植物的子葉分為子葉鞘(即基鞘)、細長的子葉聯結和棒狀的吸器3個部分,細長的子葉聯結下彎,真葉披針形而具弧形脈或劍形而具平行脈;玉蟬花幼苗子葉的基鞘和舌狀鞘均發達,子葉聯結下彎與子葉鞘部分或完全貼合,使原來的子狀鞘變成了假舌狀鞘(包括基鞘與舌狀鞘),真葉披針形;馬藺的幼苗只有舌狀鞘而基鞘完全退化,子葉聯結不但和子葉鞘完全貼合,而且還繼續伸延和下胚軸完全貼合,形成了中胚軸。以上6個種的子葉、真葉皆兩側壓扁,鞘部套褶。
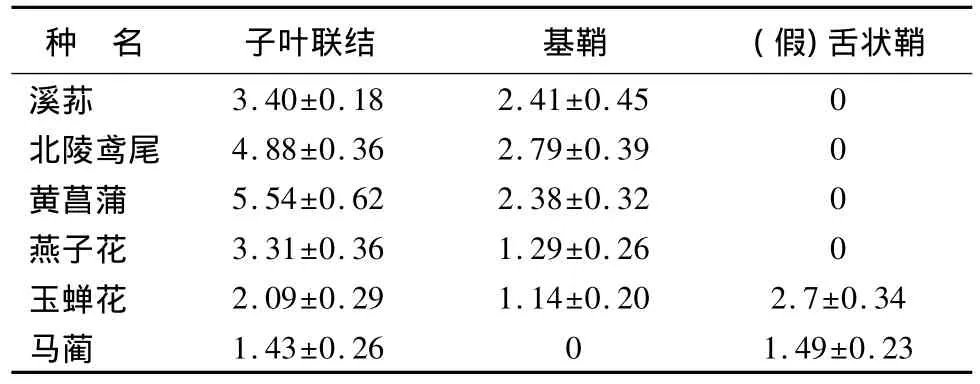
表2 6種鳶尾種苗子葉聯結及子葉鞘的長度 mm
文獻[12]將單子葉植物的幼苗類型分為21種。據此歸類,溪蓀、北陵、黃菖蒲、燕子花屬于鴨跖草型,這個類型的最大特點是細長的子葉聯結下彎;玉蟬花屬于藍耳草型,這個類型的最大特點是子葉聯結下彎和子葉鞘部分或完全貼合,使原來的子葉鞘變成了假舌狀鞘,由于貼合程度有異,子葉聯結的游離部分可位于假舌狀鞘的不同位置上;馬藺屬于迦通尼型,因為馬藺的幼苗只有舌狀鞘而基鞘完全退化,子葉聯結不但與子葉鞘完全貼合,而且還繼續伸延和下胚軸完全貼合,形成中胚軸。
4 結論與討論
從演化趨勢看,Corner[13]認為單子葉植物的種子較大并且種皮紋飾簡單的是原始特征,種子較小并且種皮紋飾復雜的為次生特征。因此可認為,鳶尾屬種皮微形態特征的網脊龍骨狀、網眼較淺的是較原始特征,表面有凹點或網脊較寬、網眼較深的是較進化的特征。由此初步推斷,鳶尾屬6種植物的進化關系是溪蓀→北陵→燕子花→黃菖蒲→玉蟬花→馬藺。
馬藺種皮表面為蜂窩狀紋飾。從形態結構上推測,這種微形態有利于它們在干旱環境下吸收和保留水分,保水力為蜂窩狀>網狀。這與前人[14-16]在其他種類上對種皮微形態的研究結論是一致的,顯示了旱生植物結構與功能的高度統一。同時也說明生態環境深刻地影響著植物形態結構的形成和差異。
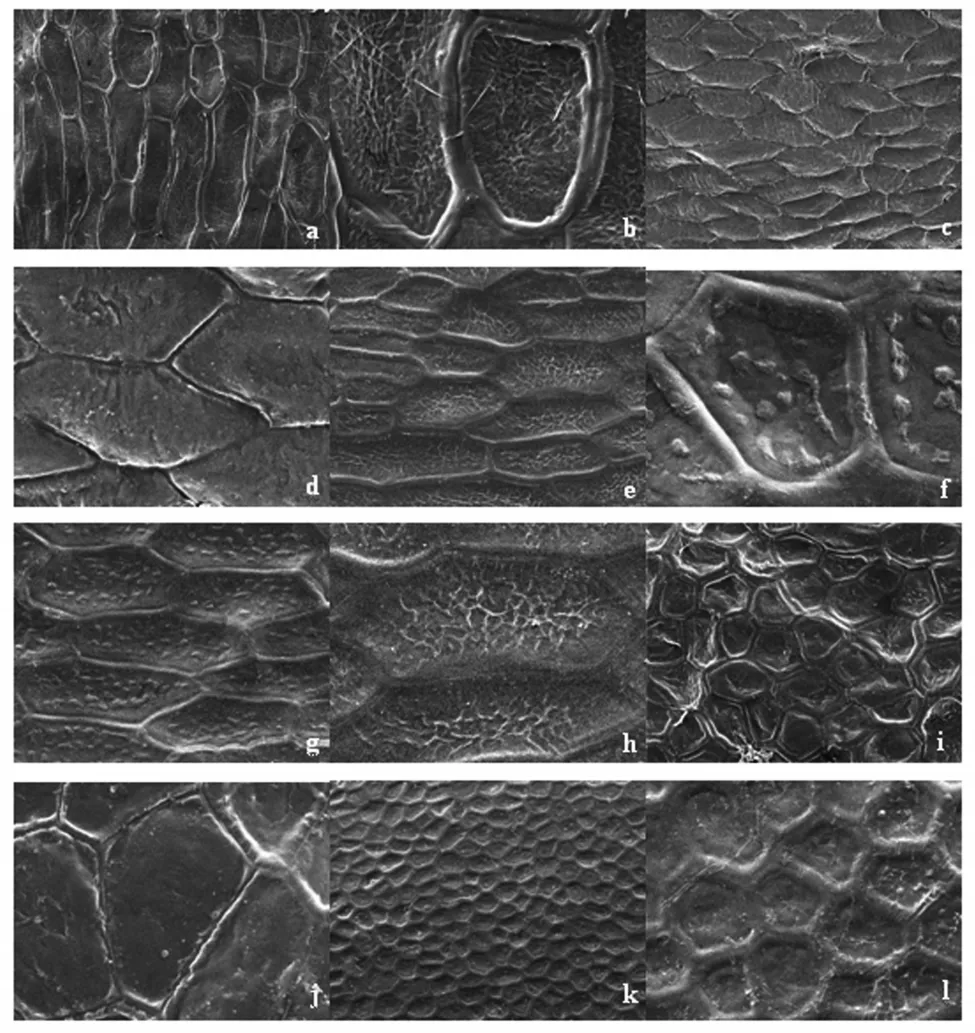
圖1 掃描電鏡下拍攝的種皮超顯微結構(300倍及1 000倍)
由表2可以看出,溪蓀、北陵、黃菖蒲、燕子花的子葉聯結較長,而玉蟬花、馬藺的子葉聯結較短。子葉鞘長度與子葉聯結長度成正比,兩者與舌狀鞘長度則成反比。Boyd[17]指出,由于舌狀鞘的存在,子葉聯結常較短,也常伴隨著子葉鞘(基鞘)的抑制。舌狀鞘比子葉鞘能更好地保護胚芽。有舌狀鞘的種類與無舌狀鞘的種類相比,由于子葉聯結短,子葉的維管束從吸器到達胚芽、胚根的聯系更加直接而經濟。因此可以推斷,有舌狀鞘的種類是較進化的類型。由此得出進化趨勢為:6種鳶尾中馬藺是最進化的種類,其次是具有舌狀鞘和基鞘的玉蟬花、黃菖蒲和燕子花,而溪蓀、北陵則是相對原始的種類。這個結論與種皮電鏡所得出的結論是一致的。
[1]Riley H P.A character analysis of colonies ofIris fulva,Iris HexagonaVar.Giganticaeruleaand natural hybrids[J].American Journal of Botany,1938,25(10):712-724.
[2]Mitra J.Karyotype analysis of bearded Iris[J].Botanical Gazette,1956,117(4):265-293.
[3]黃蘇珍,顧姻,賀善安.鳶尾屬(IrisL.)植物的雜交育種及其同工酶分析[J].植物資源與環境,1996,5(4):38-41.
[4]江明,謝文申.香根鳶尾的組織培養和快速繁殖[J].園藝學報,1995,22(3):301-302.
[5]Balkwill K,Campbell-Young G.Taxonomic studies in Acanthaceae:testa microsculpturing in southern African species ofThunbergia[J].Botanical Journal of the Linnean Society,1999,131(3):301-325.
[6]Goldblatt P.Systematics,phylogeny and evolution ofDietes(Iridaceae)[J].Annals of the Missouri Botanical Garden,1981,68(1):132-153.
[7]Manning J C,Goldblatt P.Endothecium in iridaceae and its systematic implications[J].American Journal of Botany,1990,77(4):527-532.
[8]Kron P,Stewen S C.Variability in the expession of a rhizome architecture model in a natural population ofIris versicolor(Iridaceae)[J].American Journal of Botany,1994,81(9):1128-1138.
[9]董曉東,趙毓棠,謝航.國產鳶尾屬尼泊爾鳶尾亞屬的研究[J].植物研究,1998,18(2):149-151.
[10]趙毓棠.國產鳶尾屬的研究[J].植物分類學報,1980,18(1):53-62.
[11]謝航.中國鳶尾屬有關分類群的討論及屬下分類系統的修訂[D].長春:東北師范大學,1996.
[12]葉能干,季強彪,廖海民,等.種子植物幼苗形態學[M].貴陽:貴州科學技術出版社,2002:69-87.
[13]Corner E J H.The seeds of dicotyledons[M].Cambridge:Cambridge Press,1976:161-166.
[14]孔紅.甘肅萱草屬種子微形態及其分類學意義[J].西北植物學報,2001,21(2):373-376.
[15]陳世龍,何廷農.雙蝴蝶屬和蔓龍膽屬(龍膽科)種子表面特征的研究[J].西北植物學報,2002,22(1):37-42.
[16]馬驥,王勛陵,趙松嶺.駱駝蓬屬種子微形態及其生態學與分類學意義[J].武漢植物學研究,1997,15(4):323-327.
[17]Boyd L.Monocotyledonous seeding:Morphological studies in the post seminal development of the embryo Trans[M].Proc Roy:Soc Edinb,1932:30-35.
