海濱雀稗體細胞突變體SP2008-3的特異性分析
2012-04-16 01:02:26常盼盼鐘小仙劉智微
草業(yè)學報 2012年6期
常盼盼,鐘小仙,劉智微
(1.江蘇省農(nóng)業(yè)科學院畜牧研究所,江蘇 南京210014;2.南京農(nóng)業(yè)大學動物科技學院草業(yè)科學系,江蘇 南京210095)
*海濱雀稗(Paspalumvaginatium,Seashore paspalum)為禾本科(Graminea)雀稗屬(Paspalum)植物,原產(chǎn)于美洲,分布于世界熱帶、亞熱帶,尤其美洲熱帶之海岸,為潮間帶草灘植被的主要組分,能在復雜的逆境條件下生長,其抗性主要表現(xiàn)在耐鹽、耐澇、抗旱、耐踐踏等方面,是目前已知耐鹽能力最強的草坪草種。其葉色翠綠,景觀效果優(yōu)于狗牙根(Cynodonspp.)和假儉草(Eremochloaophiuroides),已成為21世紀最具發(fā)展?jié)摿Φ呐拘筒萜翰莘N[1-7]。美國較早開展了海濱雀稗品種的選育工作,20世紀末海濱雀稗品種Adalayd從美國引進中國,1998年自深圳的觀瀾湖高爾夫球場引至江蘇省,試種結(jié)果表明,Adalayd能生長在全鹽含量為0.8%~1.0%的濱海鹽土,海水短期淹沒、較長期浸泡均能忍受,抗寒性強,綜合性狀優(yōu),在江蘇省連云港以南地區(qū)可自然越冬。鄒軼等[8]及景艷霞和袁慶華[9]系統(tǒng)研究了海鹽脅迫對海濱雀稗Adalayd生長及植株體內(nèi)陽離子含量的影響,初步明確了鹽脅迫下海濱雀稗不同于紫花苜蓿(Medicagosativa)的陽離子積累規(guī)律。鄒軼等[10]系統(tǒng)研究了海鹽脅迫對海濱雀稗Adalayd植株形態(tài)和生長的影響,并進一步明確了Adalayd對海鹽脅迫的生理響應,在此基礎上,在江蘇省和浙江省新圍墾海涂,初步建立了海濱雀稗Adalayd種莖直播和草塊直鋪技術(shù)體系,因其耐鹽性強、景觀效果好,在長江中下游及其以南地區(qū)正在備受關(guān)注。但由于海濱雀稗Adalayd與其他二倍體海濱雀稗種質(zhì)一樣,由于自交不親和[11,12],只能以無性繁殖方式擴繁,明顯影響海涂大面積草地建植以及沿海開發(fā)區(qū)大面積廠區(qū)和邊坡地綠化的成本。
截至目前,全世界只有一個商品化種子直播型海濱雀稗雜交品種“Sea Spray”(Q36313×Hyb-7),由美國純種子測試機構(gòu)(Pure Seed Testing)與喬治亞大學研究基金會(University of Georgia Research Foundation)合作培育而成[13]。據(jù)報道,2005年海濱雀稗雜交品種“Sea Spray”已正式引入我國,但尚未見其在江浙海涂推廣應用,推測可能與生態(tài)適應性和抗寒性有關(guān)。近年來,江蘇省農(nóng)業(yè)科學院鐘小仙等[14]一直致力于可自交結(jié)實型海濱雀稗新品種選育,以幼穗離體培養(yǎng)獲得的胚性愈傷組織為材料,從經(jīng)一定濃度和時間的秋水仙素誘導處理獲得的再生植株上,收獲到了體細胞突變體M1種子,M1種子均能正常發(fā)芽成苗。本研究通過比較能自交結(jié)實的海濱雀稗突變體SP2008-3和對照品種Adalayd的植物學特征和葉片氣孔特性,結(jié)合細胞遺傳學鑒定和DNA分子檢測,探索海濱雀稗突變體SP2008-3的特異性,為育成具有完全自主知識產(chǎn)權(quán)、可種子繁殖的海濱雀稗新品種提供基礎保證[15],為大面積鹽漬土利用提供技術(shù)支撐。
1 材料與方法
1.1 試驗材料
海濱雀稗品種Adalayd,2n=20,為自交不親和二倍體,20世紀末從美國夏威夷引進我國,2006年江蘇省農(nóng)業(yè)科學院從杭州白云草業(yè)研究所有限公司引進;海濱雀稗突變體SP2008-3,以Adalayd幼穗誘導產(chǎn)生的顆粒狀愈傷組織為材料[16],經(jīng)秋水仙素誘導獲得了再生植株,2009年,經(jīng)田間結(jié)實性鑒定,獲得了可自交結(jié)實的再生植株并收獲到種子,2010年4月播種出苗后植株成坪。
1.2 試驗方法
1.2.1 植物學特征的觀測 每種材料分別選取20個匍匐莖,測量從頂端向后第4個節(jié)的節(jié)間長度及直徑;選取20個直立莖,測量中間1個節(jié)的節(jié)間長度及直徑;選取30片新出的第3片完全展開葉,測量葉片長度和寬度。
1.2.2 葉片氣孔特性的觀測 分別選取2種供試材料新出的第3片完全展開葉中間部位長1 cm、寬為葉片自然寬度的部分葉片,采用離析法[17],浸泡在等量的30%過氧化氫-醋酸溶液中,在60℃烘箱內(nèi)放置24 h,待葉肉組織和表皮細胞分離后,把離析材料取出,放入裝有適量蒸餾水的培養(yǎng)皿中,用鑷子分離葉片上、下表皮,1%番紅染色1~2 min,制成臨時裝片,在OLYMPUS CX31光學顯微鏡下觀察和拍照。
采用形態(tài)學圖像分析系統(tǒng)(JD801,江蘇捷達軟件工程有限公司,南京)測定氣孔的密度、寬度和長度。每種供試材料選擇30個視野拍照,統(tǒng)計氣孔器數(shù)目,計算其密度。每種供試材料拍攝15個視野,統(tǒng)計氣孔器長度和寬度。氣孔器長度是指氣孔關(guān)閉狀態(tài)下啞鈴形氣孔器的長度;氣孔器寬度是氣孔關(guān)閉狀態(tài)下垂直于啞鈴形氣孔器的最寬值[18]。
1.2.3 細胞學鑒定 分別取海濱雀稗SP2008-3和Adalayd莖節(jié),將其放入1/2 Hoagland營養(yǎng)液中培養(yǎng)生根,待根長至1~2 cm時,于上午9:00前后取出切下根尖,經(jīng)自來水沖洗后加入到裝有預處理液8-羥基喹啉的離心管中,置于4℃冰箱中處理2~4 h。材料預處理后用自來水沖洗30 min后用卡諾氏固定液于冰箱中固定24 h,固定后的材料用蒸餾水沖洗后放入裝有預熱的1 mol/L鹽酸的離心管中于60℃恒溫水浴中解離4~6 min,解離后的材料用蒸餾水沖洗30 min后,置于卡寶品紅中染色30 min。染色后滴1滴45%冰醋酸于載玻片壓片觀察,在顯微鏡下鏡檢。在實際操作中,不同學者所統(tǒng)計細胞的數(shù)目有很大差別,一般認為85%以上的細胞具有相同染色體數(shù)時,方能確定該植物染色體數(shù)[19,20]。本試驗選擇25個染色體分散良好、形態(tài)清晰的中期分裂相細胞進行觀察、拍照和統(tǒng)計染色體數(shù)目。
1.2.4 RAPD分子鑒定 取海濱雀稗體細胞突變體SP2008-3、原始對照Adalayd(CK1)及其幼穗離體培養(yǎng)再生植株(CK2)的幼嫩葉片,采用Karroten公司生產(chǎn)的植物DNA提取試劑盒(Karroten TM Plant Genomic DNA Extraction kit)提取DNA。選取RAPD隨機引物34條委托上海生工生物工程有限公司合成。擴增反應在Ta KaRa公司PCR Thermal Cycler D上進行,反應體積25μL,內(nèi)含10×Buffer(含 Mg2+),10μmol/L d NTP,Taq DNA聚合酶0.3μL,引物0.5μmol/L,總DNA模板1μL。PCR反應程序為:94℃預變性4 min;94℃變性1 min,36℃退火1 min,72℃延伸1.5 min,共45個循環(huán),最后再72℃ 延伸7 min,4℃ 保存。擴增產(chǎn)物用1.4%的瓊脂糖凝膠電泳分離,talon全自動凝膠成像分析系統(tǒng)觀察并記錄結(jié)果。對表現(xiàn)多態(tài)性的引物,重復3次,選擇重復性好的引物進行分析。
隨機選用34個合成引物對海濱雀稗突變體SP2008-3、對照Adalayd(CK1)和Adalayd幼穗離體培養(yǎng)再生植株(CK2)進行了RAPD-PCR反應體系擴增,篩選出2個多態(tài)性和重復性好的引物S369和S131。S369序列為5′CCCTACCGAC3′;S131序列為5′TTGGTACCCC3′。
1.3 統(tǒng)計分析
采用SPSS 13.0軟件進行差異顯著性分析。
2 結(jié)果與分析
2.1 植物學特征的比較
與Adalayd相比,SP2008-3匍匐莖節(jié)間長度及匍匐莖直徑分別減小34.77%和11.76%,差異顯著(P<0.05);葉片長度及寬度分別顯著增加22.55%和11.11%;直立莖縮短變粗,但與Adalayd無顯著差異(P>0.05)(表1)。
2.2 細胞學鑒定
在統(tǒng)計的25個染色體分散良好、形態(tài)清晰的中期分裂相細胞中,SP2008-3中有22個細胞的染色體數(shù)目為20條,占所統(tǒng)計細胞總數(shù)的88%,比例超過了85%,據(jù)此確定海濱雀稗SP2008-3的體細胞染色體數(shù)目為2n=20(圖1)。

表1 SP2008-3與Adalayd植物學特征比較Table 1 Comparison on phytological characteristics between SP2008-3 and Adalayd
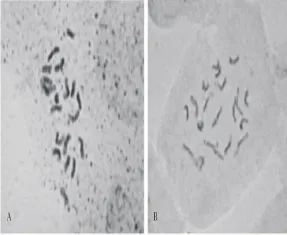
圖1 海濱雀稗突變體2008-3(A)與Adalayd(B)有絲分裂中期染色體Fig.1 Chromosome of mutant SP2008-3(A)and Adalayd(B)at mitotic metaphase
2.3 RAPD分子鑒定
引物S369序列為5′CCCTACCGAC3′,與Adalayd(CK1、CK2)相比,SP2008-3缺少1條約3 000 bp的條帶和1條約800 bp的條帶;引物S131序列為5′TTGGTACCCC3′,與 Adalayd(CK1、CK2)相比,SP2008-3植株缺少1條約200 bp的條帶(DNA Marker 5 000 bp)。再用重復性較好的引物S369對所有3個材料進行PCR擴增,不僅條帶清晰,而且經(jīng)3次重復表現(xiàn)穩(wěn)定。RAPD分析結(jié)果顯示,對照材料海濱雀稗Adalayd(CK1、CK2)均含有約1 100 bp的條帶,而SP2008-3則缺失(圖2)。
2.4 葉片表皮氣孔特性的比較
與Adalayd相比,海濱雀稗體細胞突變體SP2008-3葉片上表皮氣孔密度增加20.58%,氣孔器長度減小9.64%,氣孔器寬度減小6.31%,差異均達顯著水平。海濱雀稗體細胞突變體SP2008-3葉片下表皮氣孔器密度比Adalayd增加40.62%,氣孔器長度和氣孔器寬度分別比Adalayd減小6.73%和5.33%,且均呈顯著性差異(表2)。
3 討論
形態(tài)學鑒定是比較傳統(tǒng)、常規(guī)的品種鑒定技術(shù)[21]。劉建秀等[22]根據(jù)狗牙根匍匐莖節(jié)間長和節(jié)間直徑、葉長和葉寬等19個外部性狀對華東地區(qū)34個點上的88份狗牙根材料進行了遺傳多樣性的研究,結(jié)果表明,華東地區(qū)狗牙根可分為5個類型,即矮細型、矮粗型、斜細型、斜粗型和直立型。金洪等[23]通過對結(jié)縷草(Zoysiajaponica)葉寬、匍匐莖中部節(jié)間長、匍匐莖的直徑、穗長等表型性狀研究了結(jié)縷草居群內(nèi)部的變異類型,將20個居群分為8類。本研究通過對海濱雀稗體細胞突變體SP2008-3和Adalayd植物學特性比較發(fā)現(xiàn),SP2008-3除了可自交結(jié)實的顯著特性外,匍匐莖節(jié)間長度、匍匐莖莖粗均較Adalayd顯著減小;葉片長度和寬度均顯著增加。這表明,經(jīng)秋水仙素誘導后的SP2008-3在表型性狀上與對照材料Adalayd存在顯著差異。
葉片是進行光合作用、呼吸作用、蒸騰作用等生理代謝的主要器官,葉解剖結(jié)構(gòu)與植物生長相關(guān)性密切,因此,葉解剖結(jié)構(gòu)可作為良種篩選的又一輔助指標[24,25]。氣孔在調(diào)控水分丟失和光合作用之間總是處于一種折中狀態(tài)[26],氣孔器密度的增加有利于提高凈光合速率和品種耐濕能力,氣孔器減小則有利于減小葉片蒸騰速率[25,27,28]。與對照品種Adalayd相比,海濱雀稗突變體SP2008-3葉片上、下表皮氣孔器密度顯著增加,氣孔器長度,氣孔器寬度均顯著減小。氣孔器特性改變與生理代謝的相關(guān)性正在進一步研究。
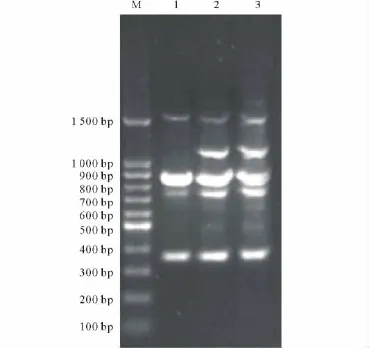
圖2 突變體SP2008-3 RAPD多態(tài)性分析Fig.2 RAPD analyses of mutant SP2008-3

表2 SP2008-3與Adalayd氣孔器特性的比較Table 2 Comparison of stomata characteristic between SP2008-3 and Adalayd
在植物誘變育種過程中,秋水仙素作為一種常用的藥劑,因作用時期不同,誘變機理和結(jié)果不同,適宜濃度的秋水仙素能破壞紡錘絲的形成,使分生細胞復制的染色體在細胞分裂時不能分向兩極,從而導致新生細胞的染色體加倍[29];而秋水仙素的作用時期為DNA復制期,其通過植物細胞在復制轉(zhuǎn)錄過程中滲入細胞內(nèi),發(fā)生堿基的替代或嵌入堿基之間,造成染色體變異,進而發(fā)生復制或轉(zhuǎn)錄錯誤,從而導致基因突變[30]。對海濱雀稗SP2008-3體細胞染色體初步計數(shù)結(jié)果表明,海濱雀稗突變體SP2008-3的體細胞染色體數(shù)目與對照品種Adalayd相同,為2n=20,但是其染色體大小、隨體著絲點位置等是否不同于Adalayd,又或者其在細胞水平上未發(fā)生改變[31,32],尚有待于進一步研究。
隨著生物實驗技術(shù)的迅速發(fā)展,植物品種鑒定的方法已從簡單、粗放的形態(tài)學鑒定進入到了分子生物學鑒定水平。常見的分子標記技術(shù)主要有RFLP、RAPD、AFLP、SSR、ISSR等[33]。其中RAPD具有操作簡便快速,對DNA的純度要求不高,DNA用量少,檢測位點多,靈敏度高等優(yōu)點[31,33]。解新明等[34]利用RAPD技術(shù)將來自廣東省海濱雀稗的3個野生居群和1個草坪型栽培品種分為兩類。本研究中海濱雀稗SP2008-3與對照Adalayd相比,缺失了1條約1 100 bp的條帶,進一步表明在DNA水平上海濱雀稗SP2008-3發(fā)生了變異。這一結(jié)果與Martelotto等[35]對2個海濱雀稗種的同源多倍體RAPD檢測結(jié)果相似,基因重組造成條帶缺失的頻率顯著高于獲得新條帶的概率。
[1] 解新明,盧小良.海雀稗種質(zhì)資源的優(yōu)良特性及其利用價值[J].華南農(nóng)業(yè)大學學報,2004,25:64-67.
[2] Duncan R R,Carrow R N.SeashorePaspalum-the Environmental Turfgrass[M].Hoboken:John Wiley&Sons,Inc.,1999.
[3] Peacock C H,Dudeck A E.Physiological and growth responses of seashore paspalum to salinity[J].HortScience,1985,20(1):111-112.
[4] Duncan R R.Seashore paspalum may be grass for the year 2000[J].Southern Turf Management,1994,5:31-32.
[5] Huang B,Duncan R R,Carrow R N.Drought-resistance mechanisms of seven warn-season turfgrass under surface soil drying:I.Shoot response[J].Crop Science,1997,37(6):1858-1863.
[6] 賀小霞,劉一明,王兆龍.海濱雀稗栽培品種的形態(tài)特征與AFLP分子標記分析[J].草地學報,2011,19(1):164-170.
[7] 鄭軼琦,臧國長,郭海林,等.假儉草雜交后代生殖性狀遺傳及相關(guān)性分析[J].草業(yè)學報,2011,20(2):283-289.
[8] 鄒軼,顧洪如,鐘小仙,等.海鹽脅迫對海濱雀稗生長及植株體內(nèi)陽離子含量的影響[J].草業(yè)科學,2009,26(4):117-120.
[9] 景艷霞,袁慶華.NaCl脅迫對苜蓿幼苗生長及不同器官中鹽離子分布的影響[J].草業(yè)學報,2011,20(2):134-139.
[10] 鄒軼,顧洪如,鐘小仙,等.海鹽脅迫對海濱雀稗生長及植株體內(nèi)陽離子含量的影響[J].草業(yè)科學,2009,24(6):117-120.
[11] Espinoza F,Quarin C L.Cytoembryology ofPaspalumchaseanumand sexual diploid biotypes of two apomicticPaspalumspecies[J].Australian Journal of Botany,1997,45:871-877.
[12] Carpenter J A.Production and use of seed in seashore paspalum[J].Australian Institute of Agricultural Science and Technology,1958,24:252-256.
[13] Fricker C,Wipff J K,Duncan R R.Hybrid seashore paspalum available from seed called“Sea Spray”[P].United States Patent:7262341B1,2007-08-28.
[14] 鐘小仙,吳娟子,劉智微,等.自交不結(jié)實海雀稗秋水仙素誘導體細胞突變體的篩選方法[P].中國專利:201110074147.1,2011-03-28.
[15] 馬金星,張吉宇,單麗燕,等.中國草品種審定登記工作進展[J].草業(yè)學報,2011,20(1):206-213.
[16] 梁流芳,佘建明,吳瑛瑛,等.海雀稗幼穗離體培養(yǎng)植株再生[J].草地學報,2008,16(6):590-593.
[17] 孫同興,江幸山.簡便有效的葉表皮離析方法——過氧化氫-醋酸法[J].廣西植物,2009,29(1):44-47.
[18] 傅志強,黃璜,何保良,等.水稻葉片氣孔特性及其相關(guān)性[J].湖南農(nóng)業(yè)大學學報:自然科學版,2007,33(6):646-650.
[19] 虞道耿,劉國道,白昌軍,等.熱研11號黑籽雀稗染色體核型分析[J].草業(yè)科學,2010,27(1):109-113.
[20] 李懋學,張贊平.作物染色體及其研究技術(shù)[M].北京:中國農(nóng)業(yè)出版社,1996:1-60.
[21] 張建成,江玉萍,王傳堂,等.花生品種鑒定技術(shù)研究進展[J].花生學報,2006,35(2):24-28.
[22] 劉建秀,賀善安,劉永東,等.華東地區(qū)狗牙根形態(tài)分類及其坪用價值[J].植物資源與環(huán)境,1996,5(3):18-22.
[23] 金洪,韓烈保,張永明.中國結(jié)縷草居群形態(tài)變異分析[J].中國草地,2004,26(2):50-56.
[24] 覃秀菊,李鳳英,何建棟,等.廣西茶樹新品種品系葉片解剖結(jié)構(gòu)特征與特性關(guān)系的研究[J].中國農(nóng)學通報,2009,25(10):36-39.
[25] 李宗艷,肖娟,蒙進芳,等.麗江牡丹和中原牡丹葉片結(jié)構(gòu)微形態(tài)比較[J].浙江農(nóng)林大學學報,2011,28(1):115-120.
[26] Casson S A,Hetherington A M.Evironmental regulation of stomatal development[J].Current Opinion in Plant Biology,2010,13(1):90-95.
[27] Aschan G,Pfanz H,Vodnik D,etal.Photosynthetic performance of vegetative and reproductive structures of green hellebore(HelleborusviridisL.agg.)[J].Photosynthetica,2005,43(1):55-64.
[28] 游明安,蓋鈞鎰,馬育華.田間條件下大豆氣孔特性的初步研究[J].大豆科學,1992,11(2):152-158.
[29] 鄭永強,徐坤.秋水仙素在植物體細胞染色體加倍中的應用研究進展[J].中國農(nóng)學通報,2003,19(5):89-91.
[30] 徐明,路鐵剛.植物誘變技術(shù)的研究進展[J].生物技術(shù)進展,2011,1(2):90-97.
[31] 甘玲,湯浩茹,董曉莉.梨種類和品種鑒定研究進展[J].中國農(nóng)學通報,2006,22(5):302-307.
[32] 陳智勇,胡忠紅,蔣建雄,等.早熟禾族3種基因型草坪草的染色體核型分析[J].草業(yè)學報,2010,19(6):281-285.
[33] 李志勇,孫啟忠,李鴻雁,等.分子標記技術(shù)在牧草種質(zhì)資源研究中的應用[J].草原與草坪,2010,30(5):91-96.
[34] 解新明,盧小良,孫雄松,等.海雀稗種質(zhì)資源分子標記的遺傳多樣性研究[J].華南農(nóng)業(yè)大學學報,2004,25:10-15.
[35] Martelotto L G,Ortiz J P A,Stein J,etal.Genome rearrangements derived from autopolyploidization inPaspalumsp.[J].Plant Science,2007,172:970-977.
