不同沙土配比對約書亞樹幼苗生理特征的影響1)
2012-03-05 09:58:00劉力寧滿秀玲唐中華曲弨琪段亮亮
東北林業大學學報 2012年7期
關鍵詞:植物
劉力寧 滿秀玲 唐中華 李 奕 曲弨琪 段亮亮
(東北林業大學,哈爾濱,150040)
在沙生環境中,幼苗的生長受到多種因素的影響[1]。土壤水分狀況是影響其生長的關鍵因素之一[2]。同一土壤吸力條件下,土壤含水量隨含沙量的增加而逐漸降低[3]。水分不僅是植物體內重要的物質組成部分,同時植物諸多生理過程均需要水分參與[4]。植物適應環境特征的研究即植物生理生態學的研究對于闡明植物適應逆境的生理生態機制,尋找植物新的抗逆途徑,從而對開發優良種質資源、育種、生態恢復以及抗旱性能評價等都具有重要意義[5]。
約書亞樹(Yucca brevifolia)是百合科(Liliaceae)絲蘭屬(Yucca L.)植物,原產于美國西南部干旱半干旱地區,廣泛分布在莫哈維沙漠[6],常綠喬木,樹形優美,耐干旱瘠薄,抗干擾,能抵抗沙漠火災[7],壽命長,在嚴酷的沙漠環境中可生存200 a 以上。它的耐熱性極強,可以忍受50 ℃以上的高溫及較大溫差,對水的需求較小,是理想的固沙、景觀植物。因此,本文選擇引進樹種約書亞樹幼苗為研究對象,在土壤不同沙土配比條件下,測定2年生約書亞樹幼苗生理生態指標的變化特征,揭示不同土壤條件下約書亞樹幼苗生理響應機制,為約書亞樹在我國成功引種奠定基礎,為我國沙區造林及荒漠化防治提供理論依據。
1 試驗材料
試驗選用長勢良好、無病蟲害的2年生約書亞樹幼苗作為試驗材料。于2010年4月20日將幼苗移栽到17 cm×18 cm 塑料營養缽中。試驗用土壤取自東北林業大學帽兒山實驗林場的自然土壤,屬腐殖質土。試驗用沙從河北省石家莊市平山縣購買,為經過篩分的天然河沙,類型為中沙。栽植土壤按體積比設計5個處理,即V(腐殖質)∶V(河沙)=9 ∶1、7 ∶3、5 ∶5、3 ∶7、1 ∶9,分別記為處理Ⅰ、處理Ⅱ、處理Ⅲ、處理Ⅳ和處理Ⅴ,每個處理有20 個重復,在溫室中進行培養,每隔30 d 澆一次水,澆灌后保證每種處理的土壤水分均達到飽和狀態。于2010年9月至10月測定約書亞樹幼苗的各項指標。利用PSYPRO 露點水勢儀測定不同處理土壤基質水勢,于澆水25 d 后測得的處理Ⅰ、處理Ⅱ、處理Ⅲ、處理Ⅳ和處理Ⅴ土壤基質水勢分別為:-42.681、-69.298、-100.191、-127.277、-151.787 kPa。
2 研究方法
2.1 光合作用指標測定
采用LI—6400 型便攜式植物光合作用儀,統一取植株中部的葉片,對各處理約書亞樹幼苗的凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)和胞間CO2的摩爾分數(Ci)[8]進行測定。測定時間為2 010年9月20日 至25日(連 續5日 晴 天),每 天8:00—16:00,每隔2 h 測定1 次。每個處理均取5個葉片進行測定,結果取平均值。
2.2 抗氧化酶及滲透調節物質測定
于2010年9月27日取幼苗葉片中間部位進行各項指標的測定,每個處理重復3 次,結果取平均值。其中:超氧化物歧化酶(SOD)活性采用硝基四唑氮藍(NBT)還原法[9];過氧化物酶(POD)活性采用愈創木酚比色法[10];過氧化氫酶(CAT)活性的測定根據Aebi 的方法[11];抗壞血酸過氧化物酶(APX)活性及谷胱甘肽還原酶(GR)活性測定參照文獻[12]的方法;蛋白質(Pr)質量分數采用考馬斯亮藍G-250 法[13];脯氨酸(Pro)采用酸性茚三酮比色法[14]。
2.3 數據處理及統計
根據測定的生理指標,檢驗約書亞樹幼苗水分利用效率、氣體交換特性在各個處理間的差異顯著性。分別計算不同處理條件下樹種的凈光合速率及蒸騰速率的日平均值[15],然后使用Excel 及SPSS多元統計分析軟件對數據進行分析。
3 結果與分析
3.1 約書亞樹幼苗生境環境因子的日變化
環境因子日變化為光合日變化測定時溫室內環境的變化。由表1可知,一天之內的光合有效輻射(PA,R)呈先升高后降低的變化趨勢。8:0 0—12:00PA,R逐漸增強。其中,10:00—12:00 增強較迅速,12:00 達到最大值(314.72 μmol·m-2·s-1),之后逐漸下降,到16:00 降至最低值(42.09 μmol·m-2·s-1)。空氣溫度(ta)的變化趨勢為先升高,持一定時間后,略微降低。由于溫室環境的影響,ta的日變化幅度較小,為21.2 ~30.0 ℃,從8:00 開始逐漸上升,12:00 達到最高,之后緩慢下降,至16:00下降為25.5 ℃,但仍高于初始溫度。空氣相對濕度(RH)先降低后稍微升高,與PA,R和ta的變化趨勢相反。8:00RH值最高(19.8%),隨后下降,到12:00 時降到最低值(10.4%),僅為最大值的1/2,之后稍微回升。空氣CO2摩爾分數(Ca)的日變化規律呈逐漸緩慢升高的趨勢。一天中溫室內Ca值在379.5 ~384.4 μmol/mol 之間,變化幅度非常小。

表1 環境因子的日變化
3.2 不同沙土配比對約書亞樹幼苗凈光合速率(Pn)及蒸騰速率(Tr)的影響
由表2可知,不同沙土配比條件下約書亞樹幼苗凈光合速率有不同的日變化規律。處理Ⅰ凈光合速率的日變化符合一般植物的光合規律,即上午10:00Pn最高,12:00 明顯降低,呈現光合“午休”現象,此后升高,下午16:00 降為最低。除處理Ⅰ外,其他處理均表現為上午8:00Pn最高,下午16:00 降為這一時段的最低值。其中處理Ⅱ上午8:00 時Pn達到6. 245 μmol·m-2·s-1,其他處理則在3 ~5 μmol·m-2·s-1范圍內變化,各處理間差異極顯著(P<0.01)。處理Ⅱ幼苗日平均Pn值最大(3.488 μmol·m-2·s-1),而 處 理Ⅳ日 平 均Pn值 較 小(1.441 μmol·m-2·s-1),二者相差2.4 倍。不同處理間Pn在不同時段差異不同,從8:00 至16:00,隨著時間的推移,各處理間差異性變小。上午8:00 左右,處理ⅡPn明顯高于其它處理,其它各處理的Pn較相近;處理Ⅰ、Ⅱ在10:00 附近Pn趨近,是其它處理Pn的3 倍多,隨著時間的推移,各處理的Pn逐漸接近,14:00 出現趨同現象;直至下午16:00,幼苗Pn出現負值,Pn伴隨著胞間CO2(Ci)摩爾分數的上升而下降,且Ci上升幅度越大,凈光合速率的下降越大(表3)。由此推斷,當溫度下降、光強減弱時,氣孔僅為次要因素,而其他非氣孔因素則成為主要因素[16]。

表2 不同處理對約書亞樹幼苗Pn 的影響

表3 不同處理下約書亞樹幼苗Ci日變化
從表4可以看出,不同時段各處理間蒸騰速率(Tr)差異很大。上午8:00 至下午14:00,各處理間Tr波動性較大,規律性差,但到下午16:00,各處理幼苗Tr較為接近。處理Ⅱ約書亞樹幼苗日平均Tr最大(1.0173 μmol·m-2·s-1),處理Ⅳ日平均Tr最小(0.4128 μmol·m-2·s-1),日平均Tr最大值是最小值的近2.5 倍。不同處理間Tr上午8:00 及中午12:00 時差異性顯著(P<0.01)。處理Ⅱ和處理Ⅴ的Tr明顯高于其它處理;處理Ⅰ、Ⅱ及Ⅳ的Tr日變化規律相似,在12:00 時蒸騰速率有明顯的下降趨勢;處理Ⅲ和處理Ⅴ的Tr日變化規律較接近,與其它處理差異顯著,即12:00 時蒸騰速率出現上升趨勢。

表4 不同處理對約書亞樹幼苗Tr 的影響
由表5可知,處理Ⅱ幼苗氣孔導度(Gs)在8:00左右明顯高于其它處理,隨著時間的推移,各處理的Gs逐漸接近,直至下午16:00,不同處理間的Gs相近。由此可見,約書亞樹幼苗Pn與Gs的變化趨勢具有同步性,說明Gs的下降是造成Pn下降的主要原因,同時也是使Tr變化的原因之一。凈光合速率下降伴隨著蒸騰速率和胞間CO2的摩爾分數下降,主要是氣孔因素,如果凈光合速率下降伴隨著胞間CO2摩爾分數的上升,則說明凈光合速率的下降以非氣孔因素為主,即凈光合速率的下降是由于葉肉細胞光合活性下降造成的[16]。約書亞樹幼苗在不同處理中Pn與Gs同步下降,凈光合速率的下降受到氣孔因素的影響,但同時伴隨著胞間CO2摩爾分數Ci的上升,這說明幼苗Pn下降既有氣孔因素,又有非氣孔因素的限制。

表5 不同處理下約書亞樹幼苗Gs日變化
3.3 不同沙土配比對約書亞樹幼苗水分利用效率(Wu,e)的影響
水分利用效率(Wu,e)是衡量植物水分消耗與物質生產之間關系的重要綜合指標[17]。本試驗用某測定時間葉片的Pn與Tr(蒸騰速率)之比表示該時刻的Wu,e[18]。由表6可見,處理Ⅰ、Ⅱ、Ⅴ水分利用率呈波動式降低,而處理Ⅲ、Ⅳ在上午10:00 Wu,e迅速升高,出現了一個峰值,分別為24. 410、17. 239 μmol·m-2·s-1,是其它處理的3 倍以上。10:00后,隨著氣溫上升,空氣相對濕度降低,為保持水分、減少蒸騰,氣孔部分關閉,導致Pn下降,但Tr仍處于較高水平[2],Wu,e逐漸降低。由于各處理14:00 左右凈光合速率的變化與蒸騰速率的變化趨勢相同,所以此時各處理的Wu,e值非常接近。Wu,e在16:00 為一天中最低值,由于此時幼苗受到非氣孔因素的影響,凈光合速率出現負值,相應的Wu,e也為負值。其中處理Ⅲ幼苗的日平均水分利用率最高,達7. 366 μmol·m-2·s-1,其次是處理Ⅳ(5.664 μmol·m-2·s-1),處理Ⅴ最低(2.990 μmol·m-2·s-1),最高值是最低值的2.5 倍。不同處理Wu,e在8:00、12:00、16:00 時都表現出極顯著差異性(P<0.01)。

表6 不同處理對約書亞樹幼苗Wu,e 的影響
3 .4 不同沙土配比對約書亞樹幼苗滲透調節物質及酶活性指標的影響
植物體內脯氨酸(Pro)是一理想的有機溶質滲透調節物[19]。脯氨酸是水溶性最大的氨基酸,被認為是有效的滲透調節物質[20-21],有助于提高植物細胞及組織的持水能力,從而增加植物抗旱性。旱生植物體內會積累大量的脯氨酸[22],脯氨酸在植物中的大量積累可能對植物的滲透調節起著重要作用[5]。由表7可知,隨著土壤中沙量的增加,約書亞樹幼苗脯氨酸質量分數表現為先增加后緩慢減少的趨勢。沙量中等的土壤,幼苗脯氨酸質量分數明顯高于沙量較低或較高的土壤。其中處理Ⅲ幼苗脯氨酸質量分數最高(4.886 μg·g-1),其次是處理Ⅳ(4.219 μg·g-1),處理Ⅰ幼苗脯氨酸的質量分數最低(1.778 μg·g-1),處理Ⅲ是處理Ⅰ的2.8 倍。不同處理間幼苗脯氨酸質量分數有較大的相關性(R2=0.853),土壤沙土配比與幼苗脯氨酸質量分數之間滿足二次曲線關系,見表8。
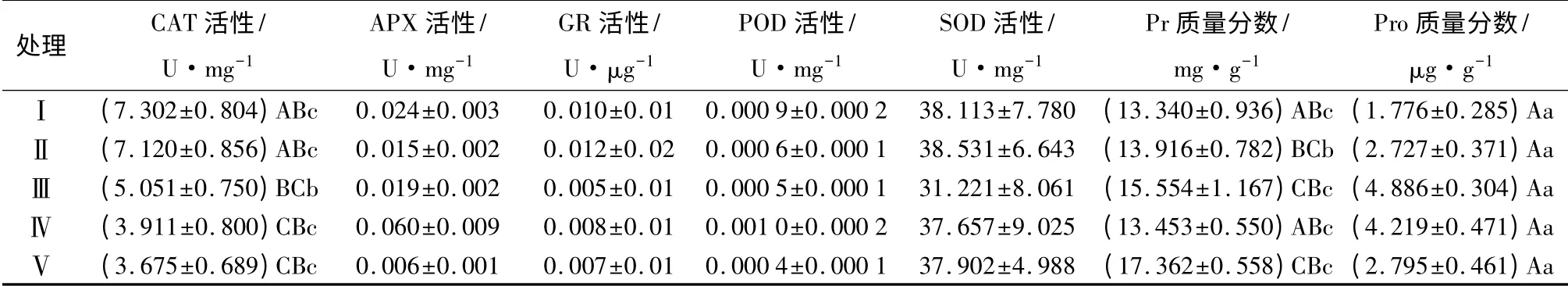
表7 不同處理下約書亞樹幼苗抗氧化指標及相關性

表8 約書亞樹幼苗生理生態指標與沙土配比的線性回歸結果
植物體內的可溶性蛋白質(Pr)質量分數是了解植物體總代謝的一個重要指標[23]。由表7可知,不同處理之間幼苗可溶性蛋白質質量分數有一定差異,處理Ⅴ幼苗可溶性蛋白質質量分數最高(17.362 mg·g-1),而處理Ⅰ可溶性蛋白質質量分數最低(13.340 mg·g-1)。不同沙土配比培育的幼苗可溶性蛋白質質量分數之間差異顯著(P<0.05)。高等植物H2O2的清除主要靠CAT、APX 等酶催化[23]。處理Ⅰ幼苗CAT 活性最強,為7.302 U·mg-1,處理Ⅴ幼苗CAT 活性最小,為3.675 U·mg-1,僅為處理Ⅰ的一半左右,不同處理間CAT 活性差異極顯著(P<0.01)。隨著土壤中沙量的增加,約書亞樹幼苗CAT 活性呈遞減趨勢,這表明土壤中沙量越少,約書亞樹幼苗體內產生的保護性酶CAT 活性越高,越有利于幼苗細胞的保護及植株的生長。另外,約書亞樹幼苗體內的APX、GR、POD 等指標的活性非常低,且不同沙土配比之間差異性不顯著(P>0.05),因此,這些酶的活性與土壤沙土配比的相關性較小(表8),但相比之下,處理Ⅲ的APX、GR、POD 及SOD 活性較其它處理均偏低。
4 結論與討論
沙量配比較高的土壤雖然導水率理想,但是對土壤的持水性也帶來負面影響。隨著沙量的增加,土壤持水性逐漸下降[3]。因此不同沙土配比的土壤環境必然會影響幼苗的生長狀況。結果表明:自然光照條件下,約書亞樹幼苗凈光合速率的變化既受氣孔因素的限制,又受到非氣孔因素的限制,但光照及溫度相對較低時非氣孔因素則成為主要因素,氣孔僅為限制其凈光合速率的次要因素。另外,不同沙土配比培育的約書亞樹幼苗光合作用存在較大差異。處理Ⅱ~Ⅳ的變化趨勢表明,隨著土壤含沙量的增加,約書亞樹幼苗的凈光合速率逐漸降低,與此同時,水分利用率呈升高趨勢[24]。造成處理Ⅰ變化趨勢不符合規律的原因,可能是約書亞樹更適宜較干旱的土壤環境,水分充足并不一定有利于其生長,而處理Ⅴ的極度缺水環境造成了幼苗生長代謝的紊亂。相比之下,在干旱的土壤環境中,處理Ⅲ的日平均水分利用率最高(7.366 μmol·m-2·s-1),而處理Ⅴ最低(2.990 μmol·m-2·s-1),僅為最高值的2/5。這說明,沙土配比5 ∶5 左右的土壤環境,可以促進約書亞樹幼苗自身光合能力的有效調節,以適應干旱的沙漠環境。
約書亞樹幼苗脯氨酸質量分數與土壤基質存在極顯著的相關關系(R2=0.853),其中處理Ⅲ幼苗的脯氨酸質量分數最高(4.886 μg·g-1),其次是處理Ⅳ(4.219 μg·g-1),而處理Ⅰ幼苗的脯氨酸質量分數最低(1.778 μg·g-1),脯氨酸質量分數最高值是最低值的2.8 倍。約書亞樹幼苗可溶性蛋白質質量分數與沙土配比之間也存在顯著的相關性(R2=0.537),其中處理Ⅴ幼苗的可溶性蛋白質質量分數最高(17.362 mg·g-1),其次是處理Ⅲ的幼苗(15.554 mg·g-1),而處理Ⅰ幼苗的可溶性蛋白質質量分數最低(13.340 mg·g-1)。研究發現:不同處理幼苗的脯氨酸質量分數、可溶性蛋白質的質量分數均與沙土配比成正相關,說明其積累的脯氨酸既具有滲透調節作用,又穩定了體內的蛋白質[5],對約書亞樹幼苗適應干旱環境具有重要作用。相對于其它處理,處理Ⅲ幼苗的脯氨酸質量分數及可溶性蛋白質質量分數均較高,說明生長在沙土配比為5 ∶5 左右的土壤環境中的約書亞樹幼苗,其體內的滲透調節物質含量較高,可以在干旱環境中更好地保護自身的細胞及組織,以適應惡劣的生長環境。另外,抗氧化酶(CAT、SOD、POD 等)活性的增強,可以加強植物體內活性氧的清除能力,從而降低膜脂的過氧化程度,保護膜結構的完整性[25]。約書亞樹幼苗CAT活性直接受沙土配比的影響,且呈顯著負相關(r=-0.886,P<0.05)。不同處理間幼苗CAT 活性差異極顯著(R2=0.925),隨著含沙量的增加,約書亞樹幼苗CAT 活性呈遞減趨勢。CAT 活性的受抑,說明體內酶性防御系統受到削弱[26]。試驗表明,配比土壤中沙量的增加會導致約書亞樹幼苗體內CAT 酶防御系統受破壞,使CAT 酶對幼苗的保護能力削弱。而約書亞樹幼苗SOD、POD、APX、GR 活性與沙土配比的相關性小,且不同處理之間差異性不顯著(P>0.05)。但相比之下,處理Ⅲ的APX、GR、POD及SOD 活性較其它處理均偏低,說明處理Ⅲ土壤生境對約書亞樹幼苗的脅迫最小,幼苗更適應這種土壤環境。
由于沙漠干旱的土壤環境會對幼苗生理生態特征產生不同程度的影響,在旱生土壤生境中,滲透調節物質含量增高,可以有效提高約書亞樹幼苗的抗性能力。綜上所述,幼苗在沙土配比為5 ∶5 左右的土壤中生長,無論是水分利用率、脯氨酸質量分數、可溶性蛋白質量分數都相對較高,且抗氧化酶的活性相對較低,較其它處理對干旱的適應性較強。所以,如果在沙區栽植約書亞樹,可以在約書亞樹幼苗期利用客土的方法為其創造沙土配比為5 ∶5 左右的土壤小生境,以更好地適應惡劣、干旱的生存環境,從而有效地促進荒漠地區約書亞樹幼苗的生長。
[1] 李文婷,張超,王飛.沙埋與供水對毛烏素沙地兩種重要沙生植物幼苗生長的影響[J].生態學報,2010,30(5):1192-1199.
[2] 姬蘭柱,肖冬梅,王淼. 模擬水分脅迫對水曲柳光合速率及水分利用效率的影響[J].應用生態學報,2005,16(3):408-412.
[3] 宋桂龍,韓烈保,李德穎.不同沙土配比根系層土壤的持水特性研究[J].土壤通報,2008,29(2):233-237.
[4] 鄭明清,鄭元潤,姜聯合.毛烏素沙地4 種沙生植物種子萌發及出苗對沙埋及單次供水的響應[J].生態學報,2006,26(8):2474-2484.
[5] 胡小文,王彥榮,武艷培.荒漠草原植物抗旱生理生態學研究進展[J].草業學報,2004,13(3):10-15.
[6] Loik M E,Huxman T E,Hamerlynck E P,et al. Low temperature tolerance and cold acclimation for seedlings of three Mojave Desert Yucca species exposed to elevated CO2[J]. Journal of Arid Environments,2000,46(1):43-56.
[7] DeFalco L A,Esque T C,Scoles-Sciulla S J,et al. Desert wildfire and severe drought diminish survivorship of the long-lived Joshua tree (Yucca brevifolia;Agavaceae)[J]. American Journal of Botany,2010,97(2):243-250.
[8] Nijs I,Ferris R,Blum H. Stomatal regulation in a changing climate:a field study using Free Air Temperature Increase (FATI)and Free Air CO2Enrichment (FACE)[J]. Plant,Cell and Environment,1997,20(8):1041-1050.
[9] Krivosheeva A,Tao Dali,Ottander C,et al. Cold acclimation and photoinhibition of photosynthesis in Scots pine[J]. Planta,1996,200(3):296-305.
[10] 張以順,黃霞,陳云鳳.植物生理學實驗教程[M].北京:高等教育出版社,2009:139-141.
[11] Aebi H. Catalase in vitro[J]. Methods in Enzymology,1984,105(13):121-126.
[12] Ma Fengwang,Cheng Lailiang. The sun-exposed peel of apple fruit has higher xanthophyll cycledependent thermal dissipation and antioxidants of the ascorbate glutathione pathway than the shaded peel[J]. Plant Science,2003,165(4):819-827.
[13] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry,1976,72:248-254.
[14] 趙樹蘭,廉菲,多立安.草坪植物對生活垃圾堆肥基質的生理生態響應[J].生態學報,2009,29(2):916-923.
[15] 徐利霞,楊水平,姚小華.石漠化地區3 個樹種幼苗在水分脅迫下的光合特性與抗旱性關系[J]. 林業科學研究,2006,19(6):785-790.
[16] Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33:317-345.
[17] 王美云,李少昆,趙明.關于玉米光合作用與葉片水分利用效率研究中的作用[J].作物學報,1997,23(3):345-352.
[18] 張正斌,山侖.小麥旗葉水分利用效率比較研究[J]. 科學通報,1997,42(17):1876-1881.
[19] 周瑞蓮,孫國鈞,王海鷗. 沙生植物滲透調節物對干旱、高溫的響應及其在抗逆性中的作用[J].中國沙漠,1999,19(1):18-22.
[20] 馬玉心,蔡體久,宋麗萍,等. 興安鹿蹄草(Pyrola dahurica(H. Andr. )Kom.)在雪蓋前后丙二醛及滲透調節物質的變化規律[J].生態學報,2007,27(11):4596-4602.
[21] Hare P D,Cress W A,Van Staden C J. Dissecting the roles of osmolyte accumulation during stress[J]. Plant,Cell and Environment,1998,21(6):535-553.
[22] Wang Soumin,Wan Changgui,Wang Yanrong,et al. The characteristics of Na+,K+and free proline distriribution in several drought-resistant plants of the A lxa Desert,China[J]. Journal of A rid Environments,2004,56(3):525-539.
[23] Asada K. Ascorbate peroxidase—a hydrogen peroxide scavenging enzymes in plants[J]. Physiologia Plantarum,1992,85(2):235-241.
[24] 牛書麗,蔣高明.內蒙古渾善達克沙地97 種植物的光合生理特征[J].植物生態學報,2003,27(3):318-324.
[25] Agarwal S,Sairam R K,Srivastava G C,et al. Changes in antioxidant enzymes activity and oxidative stress by abscisic acid and salicylic acid in wheat genotypes[J]. Biol Plant,2005,49(4):541-550.
[26] 鈕福祥,華希新,郭小丁,等. 甘薯品種抗旱性生理指標及其綜合評價初探[J].作物學報,1996,22(4):392-398.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
