登革Ⅱ型病毒E蛋白Ⅰ、Ⅱ結構域在大腸桿菌中的表達及免疫反應性鑒定*
2012-01-24 02:12:58付宇姣鄭學禮
中國人獸共患病學報 2012年5期
付宇姣,鄭學禮
登革病毒(DENV)屬于黃病毒科黃病毒屬,是最常見的通過節肢動物(伊蚊)傳播的病毒性疾病病原體,分布于全球100多個國家,特別是熱帶和亞熱帶地區[1]。全球大約有25億人受登革病毒感染的威脅,每年大約有5 000萬人感染登革熱,其中大約有210萬的嚴重病例,50萬例登革出血熱,20 000例死亡病例。登革病毒感染引起的病變范圍很廣,從自限性疾病到嚴重危及生命的登革熱、登革出血熱以及登革休克綜合征[2-4]。登革熱/登革出血熱已成為熱帶地區最為重要的蚊-傳播病毒性疾病,是重要的公共衛生問題[5]。
登革病毒是一個正義單鏈RNA病毒。登革病毒基因組RNA編碼3個結構蛋白(C、E、prM/M)和7個非結構蛋白(NS1、NS2a、NS2b、NS3、NS4a、NS4b、NS5),分為4個典型的血清型(DENV1-4)。包膜糖蛋白E是主要的病毒顆粒包膜糖蛋白,以同源三聚體形式存在于成熟病毒顆粒表面。結晶學的研究表明,登革病毒的包膜糖蛋白E與其他黃病毒類似,分為3個結構域(Ⅰ-Ⅲ)[6-9]。在三個結構中,β鏈形成的二級結構是其主要結構。結構域I位于E蛋白單體中央,E蛋白II區折疊成長的指樣結構,E蛋白C端的氨基酸殘基構成了結構域Ⅲ。因為在所有成熟具有感染性的黃病毒中E蛋白均是由與病毒融合有關的同源二聚體形成,推斷E蛋白的所有結構都可能和病毒與細胞的融合過程有關[10]。其中E蛋白II區中第98~111位氨基酸殘基組成的cd環,富含甘氨酸且具有很強的疏水性,序列組成在幾乎所有的蟲媒黃病毒中高度保守。最近研究發現,cd環可在E蛋白由二聚體向三聚體的轉變過程中插入胞膜,參與病毒與宿主細胞膜的融合,被認為是病毒融合靶點,或認為是潛在的病毒融合肽[11-12]。推斷第一、第二結構域可能與登革病毒發病機制相關。本研究旨在從大腸桿菌中表達登革II型病毒包膜糖蛋白(E)的第一、二結構域(DI,DII),并通過免疫印跡驗證重組蛋白的免疫原性,為后續的功能研究奠定工作基礎。
1 材料與方法
1.1 實驗材料
1.1.1 病毒株、菌株和質粒 登革Ⅱ型病毒NGC株、大腸桿菌DH5ɑ、表達菌BL21(DE3)、表達載體質粒pET-28a(+)為本實驗室保存。
1.1.2 主要試劑、耗材 RT-PCR試劑盒、DNAMarker、限制性內切酶等購自大連寶生物工程有限公司;質粒提取試劑盒和凝膠回收試劑盒購自美國O-mega公司;PVDF膜購自millipore公司;Trizol試劑、His·Tag單抗、增強型HRP-DAB底物顯色試劑盒購自英韋創津公司;HRP標記的羊抗小鼠IgG購自武漢博士德生物工程有限公司;登革病毒單抗(D1-11)購自美國Santa Cruz生物技術公司;預染雙色蛋白質標準購自美國Bio-Rad公司。
1.1.3 實驗動物 實驗用1~3日齡昆明乳鼠購自南方醫科大學實驗動物中心。
1.2 方法
1.2.1 引物的合成 引物用Premier 5.0軟件設計引物并由生工生物工程(上海)有限公司合成。RT-PCR擴增E蛋白第一,二結構域(DⅠ,Ⅱ)基因片 段 的 引 物:DV2-E-F:5′-CGTCCATATGATGCGTTGCATAG-3′(下劃線部分為NdeⅠ酶切位點 ),DV2-E-R:5′-TCGCCTCGAGTTATCCTTTAGAGC-3′(下劃線部分為XhoⅠ酶切位點)。
1.2.2 乳鼠腦內接種病毒傳代 取-70℃保存的登革Ⅱ型病毒,加入DMEM培養基稀釋作為病毒液。用微量注身器每只30μL顱內接種1~3d齡昆明乳鼠。接種后5~9d期間的發病瀕死乳鼠拉頸處死,無菌取腦組織備總RNA的提取。
1.2.3 目的基因的獲得 發病乳鼠腦組織的總RNA提取參見Invitrogen公司的Trizol說明書。E蛋白第一、二結構域基因片段的擴增參見TaKa-Ra公司的RT-PCR試劑盒說明書。反應條件為:94℃預變性5min,94℃變性45s,58.1℃退火60 s,72℃延伸120s,30個循環;72℃延伸8min。反應后擴增產物經1%瓊脂糖凝膠電泳鑒定,檢測擴增片段的長度。
1.2.4 目的基因的克隆及重組表達質粒的構建將目的基因的PCR產物經切膠回收純化后與經XhoⅠ及NdeⅠ酶切純化的表達載體pET28a(+),按照TaKaRa公司T4DNA ligase試劑盒說明書連接目的片斷與表達載體,連接產物轉化DH5α感受態細胞,置37℃培養12~16h挑取單菌落,采取菌落PCR法初篩陽性克隆質粒,提取質粒并進行雙酶切鑒定,將陽性克隆質粒送上海英俊生物工程有限公司測序,檢查連入的片段有無堿基突變及開放讀碼框是否正確。測序正確的質粒命名為pET28a-DV2-E-D1&2。
1.2.5 重組質粒轉化BL21(DE3)感受態細胞及誘導表達的SDS-PAGE分析 測序正確的pET-28a(+ )-DV2-E-D1&2,pET-28a(+ ),轉 化 BL21(DE3)感受態細胞,挑取單菌落于終濃度為50mg/L卡那霉素的LB培養基中,37℃振蕩培養至OD600為0.5,取部分菌液作為誘導前對照,剩余菌液加入IPTG至終濃度為1mmol/L,37℃振蕩培養3h后收集菌液,沉淀,經12%分離膠的SDS-PAGE分析重組菌的誘導表達。
1.2.6 重組蛋白的可溶性分析及純化 菌體沉淀重懸于裂解緩沖液中冰浴超聲裂解至澄清,裂解菌液離心后,回收上清及沉淀并加入等體積的裂解緩沖液重懸沉淀,分別取等體積的上清與重懸沉淀經12%分離膠的SDS-PAGE分析重組蛋白的可溶性,重組蛋白的純化步驟參見德國Novagen公司的Ni-NTA His·Bind樹脂說明書。
1.2.7 Western blot鑒定重組蛋白誘導前后的重組菌全菌蛋白 重組蛋白經12%SDS-PAGE分離后,半干轉印于 PVDF 膜上,0.3%Tween-20,1%BSA封閉1h后,分別加入1∶3 000稀釋的His·Tag單抗和1∶500稀釋的登革病毒單抗于37℃孵育1.5h,PBST洗膜3次,加入1∶500稀釋的HRP標記的羊抗小鼠IgG于37℃孵育1h,PBST充分洗膜后按照增強型HRP-DAB底物顯色試劑盒說明書顯色。
2 結 果
2.1 以提取的乳鼠總RNA經過RT-PCR擴增得到約900bp的E蛋白第一、二結構域基因片段,見圖1。
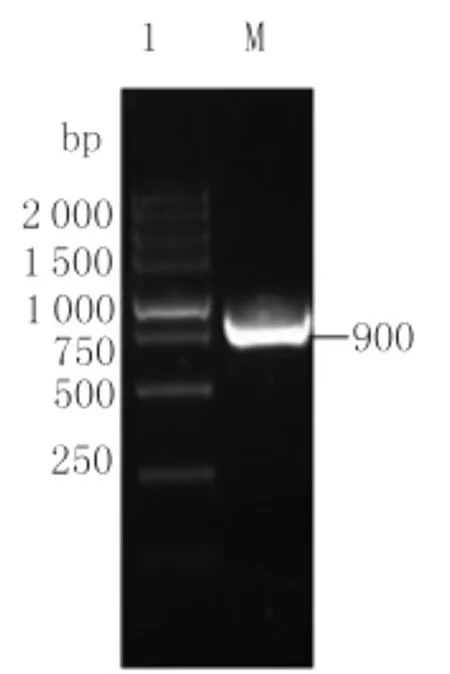
圖1 E蛋白第一,二結構域基因片段的PCR擴增Fig.1 PCR amplification of the envelop protein DNA fragmentLane 1:Products of PCR amplification M:DL-5000marker
2.2 pET28a-DV2-E-D1&2載體的構建 重組質粒pET28a-DV2-E-D1&2經XhoⅠ和 NdeⅠ雙酶切后,可見約900bp的酶切片段,與預期大小相符,見圖2。測序分析表明,插入序列正確且開放讀碼框正確。
2.3 重組菌誘導表達的SDS-PAGE分析 重組菌BL21(DE3)/pET-28a(+)-DV2-E-D1&2經IPTG誘導后在37kD位置有明顯的誘導后表達帶,其大小與預測值一致。超聲裂解重組菌后,分別取上清與沉淀經12%SDS-PAGE分析,顯示重組蛋白主要以包涵體形式存在于沉淀中,見圖3。
2.4 Western blot鑒定重組蛋白 Western blot結果顯示,重組菌誘導前的全菌蛋白沒有與His·Tag單抗反應的條帶,而誘導后的全菌蛋白和在相對分子質量為37000的位置均有與His·Tag單抗反應的特異條帶,見圖4。登革病毒單抗(D1-11)與重組誘導前的全菌蛋白無反應,只在誘導后的全菌蛋白中有特異的條帶出現,且與預測的相對分子質量為37000的位置相符合,見圖5。上述結果說明,重組蛋白具有良好的免疫反應性。

圖2 重組質粒pET-28a-DV2-E-D1&2的酶切鑒定Fig.2 Identification of the recombinant plasmid pET-28a(+)-DV2-E-D1&2by restriction enzymes digestionLane 1,3:Recombinant plasmid pET-28a(+)-DV2-E-D1&2;Lane 2,4:Recombinant plasmid pET-28a(+)-DV2-E-D1&2digested by the restriction enzymes XhoⅠ/NdeⅠ;M :DL-5000Marker
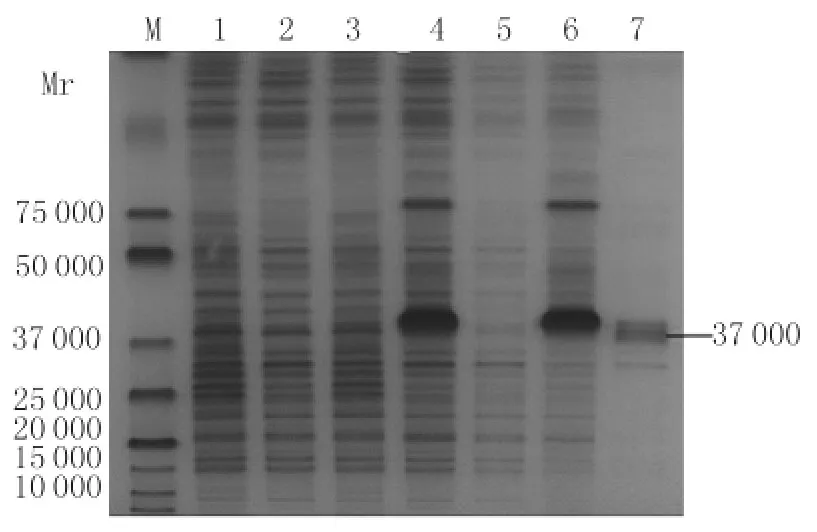
圖3 BL21(DE3)/pET-28a(+)-DV2-E-D1&2重組菌表達的 SDS-PAGE 分析Fig.3 SDS-PAGE analysis the expression of BL21(DE3)/pET-28a(+)-DV2-E-D1&2Lane 1:BL21(DE3)/pET-28a(+)before induction;Lane 2:BL21(DE3)/pET-28a(+)after induction;Lane 3:BL21(DE3)/pET-28a(+)-DV2-E-D1&2 before induction;Lane 4:BL21(DE3)/pET-28a(+)-DV2-E-D1&2 after induction;Lane 5:Supernatant of BL21(DE3)/pET-28a(+)-DV2-E-D1&2after sonication;Lane 6:Pellet of BL21(DE3)/pET-28a(+)-DV2-E-D1&2after sonication;Lane 7:Purification of the recombinant protein DV2-E-D1&2with Ni-NTAHis·Bind Resin;M:Protein marker

圖4 His·tag單克隆抗體鑒定重組DENV-2-E-D1&2蛋白Fig.4 Western Blot identification of recombinant protein with His·Tag McAbLane 1:BL21(DE3)/pET-28a(+)-DV2-E-D1&2 after induction;Lane 2:BL21(DE3)/pET-28a(+)-DV2-E-D1&2 before induction;M:Prestained Protein marker

圖5 Dengue Virus(D1-11)的單克隆抗體鑒定重組DENV-2-E-D1&2蛋白Fig.5 Western blot identification of recombinant protein with DENV(I-IV)McAbLane 1:BL21(DE3)/pET-28a(+)-DV2-E-D1&2 after induction;Lane 2:BL21(DE3)/pET-28a(+)-DV2-E-D1&2 before induction;M:Prestained Protein marker
3 討 論
登革病毒包膜蛋白E是由494~501個氨基酸殘基組成、分子質量約為55~60kDa的糖蛋白。E蛋白可形成3個不同的結構域,即呈β桶狀的I區,參與二聚體形成的II區和具有免疫球蛋白樣結構的III區。硫酸乙酰肝素(HS)是廣泛存在于細胞表面的一類由二糖重復單位組成的多糖,表面具有大量負電荷,具有包括介導病原體吸附等在內的多種生物學功能。研究表明,登革2型病毒中含高度保守的堿性氨基酸Lys89,Lys122,Lys123,其中I區和II區中帶有大量的正電荷Arg和Lys可能有助于病毒與硫酸乙酰肝素結合。研究也發現,將細胞表面的硫酸乙酰肝素脫硫或經酶消化去除后,登革病毒和黃熱病毒與細胞的結合能力顯著降低[13]。推斷第一、第二結構域可能與登革病毒發病機制相關。
本研究中重組表達的蛋白經SDS-PAGE分析以包涵體的形式存在。分析其形成原因有以下幾方面:蛋白表達量過高;重組蛋白含巰基酸多;重組蛋白所處發酵溫度高或胞內pH接近蛋白的等電點,這些因素均可導致降低蛋白的溶解度,形成包涵體。
本研究成功地在大腸桿菌中表達登革II型病毒包膜糖蛋白(E)的第一、二結構域(DI,DII),對其進行免疫反應性鑒定,鑒定結果表明其具有良好的免疫反應性。為進一步研究第一、二結構域與宿主細胞的相互作用和功能及其致病性奠定了基礎。
[1]Henchal EA,Putnak JR.The dengue viruses[J].Clin Microbiol Rev,1990,3(4):376-96.
[2]Guzman MG,Halstead SB,Artsob H,et al.Dengue a continuing global threat[J].Nat Rev Microbiol,2010,8(12Suppl):7-16.
[3]Whitehorn J,Farrar J.Dengue[J].Br Med Bull,2010,95(1):161-73.
[4]Tapia-CR,Méndez-Galván JF,Gallardo-Rincón H.The growing burden of dengue in Latin America[J].Clin Virol,2009,46(2 Suppl):3-6.
[5]雷子慶,蘇裕心,鄭學禮.登革域型病毒E蛋白第三結構域在大腸桿菌中的表達、純化及免疫反應性鑒定[J].南方醫科大學學報,2010,30(7):1496-1499.
[6]Zhang W,Chipman PR,Corver J,et al.Visualization of membrane protein domains by cryo-electron microscopyof dengue virus[J].Nat Struct Biol,2003,10(11):907–912.
[7]Kuhn RJ,ZhangW,Rossmann MG,et al.Structure of dengue virus:implications for flavivirus organization,maturation,and fusion[J].Cell,2002,108(5):717-725.
[8]Modis Y,Ogata S,Clements D,et al.Structure of the dengue virus envelope protein after membrane fusion[J].Nature 2004,427(6972):313-319.
[9]Megret F,Hugnot JP,Falconar A,et al.Use of recombinant fusion proteins and monoclonal antibodies to define linear and discontinuous antigenic sites on the dengue virus envelope glycoprotein[J].Virology,1992,187:480-491.
[10]Kuhn RJ,Zhang W,Rossman MG,et al.Structure of dengue virus:implications for flavivirus organization,maturation,and fusion[J].Cell,2002,108(5):717-725.
[11]Zheng A,Umashankar M,Kielian M.In vitro and in Vivo studies identify important features of dengue virus pr-E protein interactions[J].PLoS Pathog,2010,6(10):e1001157.
[12]Lin SR,Zou G,Hsieh SC.The helical domains of the stem region of dengue virus envelope protein are involved in both virus assembly and entry [J].Journal 0fVirology,2011,85(10):5159-5171.
[13]秦鄂德,秦成峰,姜濤.登革病毒與登革病毒病[M].北京:科學出版社,2008:28-34.
