RP-HPLC法同時測定釀酒酵母胞內磷酸腺苷和輔酶I
2012-01-09 05:09:46李會品趙謀明俞志敏雷宏杰趙海鋒
食品與生物技術學報 2012年5期
李會品, 趙謀明, 俞志敏, 雷宏杰, 趙海鋒
(華南理工大學 輕工與食品學院,廣東 廣州 510640)
RP-HPLC法同時測定釀酒酵母胞內磷酸腺苷和輔酶I
李會品, 趙謀明, 俞志敏, 雷宏杰, 趙海鋒*
(華南理工大學 輕工與食品學院,廣東 廣州 510640)
為準確監控釀酒酵母胞內磷酸腺苷和輔酶I的代謝情況,作者建立了釀酒酵母胞內磷酸腺苷和輔酶I的高效液相色譜分析方法。胞內代謝物經液氮研磨提取后,采用Agilent Zorbax SB-Aq色譜柱(5μm,4.6 mm×150 mm)分離,流動相0.025 mmol/L三乙胺-磷酸緩沖液(p H 6.0)與乙腈梯度洗脫,流速1 m L/mim,檢測波長254 nm。在此條件下,3種磷酸腺苷和兩種輔酶I達到了較好的分離效果。回收率在98.30%~103.42%之間,RSD<3.00%。結果表明,本法靈敏、準確、重現性好,可用于釀酒酵母胞內磷酸腺苷和輔酶I摩爾質量濃度的測定。
高效液相色譜;釀酒酵母;磷酸腺苷;輔酶I
啤酒作為世界性飲料,是世界上產量最大的飲 料酒[1]。啤酒釀造過程中,酵母將麥汁中的糖分發酵生成乙醇、二氧化碳和其他副產物。這一過程包括諸多代謝反應,其反應活性直接決定啤酒的品質[2]。酵母代謝活性受胞內磷酸腺苷(ATP、ADP和AMP)和輔酶I(NADH和NAD+)含量和狀態的調控[3-4]。ATP是釀酒酵母細胞合成的含有高能磷酸鍵的化合物,參與機體內蛋白質、脂肪及糖類的代謝,是生物體重要的能量物質,對生物體正常機能的維持有著無可替代的作用。ATP可水解生成ADP和AMP,三者之間的相互轉化使ATP在機體內達到動態平衡,以維持機體代謝的需要[5]。NAD+、NADH是輔酶I在細胞內存在的兩種形式。細胞代謝過程中,氧化型和還原型輔酶I之間的相互轉換,起著調節能量代謝和物質代謝的核心作用[6]。
目前,測定ATP、ADP和AMP的方法較多,主要有試劑盒法、電化學分析法、層析法、生物發光法、高效液相色譜(HPLC)法、毛細管區帶電泳法及熒光光譜法[7]。其中試劑盒法、電化學分析法、生物發光法及毛細管區帶電泳法存在操作繁瑣、精度低、雜質干擾等缺點,HPLC法則具有操作簡便、靈敏度高、精確度高等特點[8-9]。但準確分析檢測酵母胞內的磷酸腺苷和輔酶I,并監測其在發酵過程中的代謝情況,尚未見相關報道。因此,作者在以前研究的基礎上,建立了利用RP-HPLC法測定釀酒酵母胞內磷酸腺苷和輔酶I的方法,該方法不僅操作簡便,而且穩定性、重現性好,準確度、靈敏度高,能為啤酒釀造者監控釀酒酵母的代謝活性提供可靠依據。
1 材料與方法
1.1 菌株
下面釀酒酵母(Saccharomyces cerevisiae)FBY0095,上面釀酒酵母(Saccharomyces cerevisiae)FBY0099:由作者所在實驗室保藏。
1.2 儀器與試劑
Waters 600高效液相色譜儀,Waters 2418紫外檢測器,Agilent Zorbax SB-Aq色譜柱(5μm,4.6 mm×150 mm);p HS-2C型數字酸度計:上海精科儀器有限公司;3-18K高速冷凍離心機:Sigma儀器有限公司;XO-650D超聲細胞破碎儀:南京先歐生物科技有限公司;三乙胺、乙腈、磷酸:迪馬試劑有限公司,色譜純;ATP-Na2、ADP-Na2、AMPNa2、NAD+、NADH:Sigma試劑有限公司,色譜純;其他試劑均為分析純。
1.3 酵母細胞猝滅
取10 m L對數期發酵液,加入40 m L、-40℃、體積分數72%甲醇溶液,在-20℃下保藏10 min,每兩分鐘劇烈振蕩一次,以猝滅酵母胞內代謝。在4℃下以6 000 r/min離心10 min,收集菌體,再用冰甲醇溶液洗滌兩遍,離心后收集菌體[10]。
1.4 胞內物提取
1.4.1 超聲提取向菌體中加入5 m L pH 6.0的磷酸鈉緩沖液,超聲處理10 min(功率300 W,工作1 s,間歇0.5 s),離心后收集上清液[11]。
1.4.2 熱乙醇提取向菌體中加入一定量熱乙醇,在沸水浴中將乙醇蒸干,然后加入5 m L pH 6.0的磷酸鈉緩沖液,煮沸10 min,離心后收集上清液[10]。
1.4.3 液氮提取向菌體中加入2 m L pH 6.0的磷酸鈉緩沖液,用石英砂研磨5 min,再加入3 m L磷酸鈉緩沖液洗滌,離心后收集上清液,并于-20℃下保藏。
1.5 色譜條件
流動相:0.025 mmol/L三乙胺溶液(用磷酸調p H至6.0)和乙腈梯度洗脫,流速1 m L/min,洗脫時間40 min,在0~20 min,流動相中三乙胺緩沖液與乙腈的比例由99∶1降低到95∶5,保持5 min,接下來15 min內,流動相比例再由95∶5變化為99∶1,紫外檢測波長254 nm,柱溫30℃,進樣量20μL。
2 結果與分析
2.1 提取方法的選擇
釀酒酵母細胞經甲醇猝滅處理后,用不同提取方法對胞內代謝物進行提取,然后進行HPLC檢測。由表1可知,液氮研磨法對胞內磷酸腺苷和輔酶I的提取效果最好,該法提取的總磷酸腺苷和總輔酶I濃度均為最高。超聲提取法提取效果次之,熱乙醇提取法提取效果最差。這說明液氮研磨法對酵母細胞有較好的破碎作用,使胞內代謝物完全釋放出來。超聲處理對酵母細胞產生空化作用,使細胞膜產生裂隙,胞內物質不完全釋放[12]。乙醇對細胞膜起溶解作用,但酵母細胞膜較堅固,所以只有部分胞內物被提取出來。超聲提取法對ATP和NADH的水解作用較明顯,使細胞能荷(EC)和還原度(NADH/NAD+)最低,不能準確反映酵母細胞的代謝狀況。液氮研磨提取后,酵母細胞的EC和還原度均為最高,可能是由于在低溫條件下能較好地防止ATP和NADH的水解。因此,液氮研磨法對酵母胞內磷酸腺苷和輔酶I的提取效果較好。

表1 不同提取方法對釀酒酵母胞內磷酸腺苷和輔酶I提取的影響Tab.1 Effects of different methods on extraction of intracellular adenosine phosphate and coenzyme I from Saccharomyces cerevisiae(μmol/g)
2.2 流動相條件選擇
2.2.1 流動相種類選擇磷酸鹽緩沖液-乙腈和三乙胺緩沖液-乙腈系統是分離磷酸腺苷和輔酶I時應用較廣泛的流動相[13-14]。作者選擇 Na2HPO4-Na H2PO4緩沖液-乙腈和三乙胺-磷酸緩沖液-乙腈作為流動相,比較5種物質的分離情況。結果表明,這兩種流動相都能較好的分離磷酸腺苷和輔酶I,相同濃度條件下,當三乙胺緩沖液作為流動相時,磷酸腺苷和輔酶I的響應值較大。此外,由于磷酸鹽緩沖液的本底吸收較高,故選擇三乙胺緩沖液作為流動相。
緩沖液濃度直接影響緩沖能力大小,影響流動相穩定性,從而影響色譜柱分離效果。0.025、0.03、0.035、0.04 mmol/L 4種濃度的緩沖液對磷酸腺苷和輔酶I的分離效果都較好。而高濃度緩沖液會增加流動相的本底吸收,降低靈敏度,影響分離效果,并縮短色譜柱壽命。因此,選擇0.025 mmol/L的三乙胺緩沖液[15]。
2.2.2 流動相p H選擇由于流動相p H值對磷酸腺苷的保留時間有較大影響,因此考察不同流動相p H 值(p H 5.0、5.5、6.0、6.5、7.0)對磷酸腺苷和輔酶I分離的影響,結果見圖1。由圖1知,AMP、ADP、ATP和NADH的保留時間隨p H值的增加而減小,NADH的降低幅度較大,而NAD+的保留時間隨p H的增加而增加。p H值為5.0時,5種物質保留時間差異較大,較易分離,但NADH的保留時間過大對分離不利。p H 為5.5時,NAD+與 AMP不能分離開來。p H 為6.5時,NAD+與ADP的保留時間較接近,不易分離。p H為7時,5種胞內物分離效果較好,但ATP和NADH在酸性條件下穩定性較好,在堿性或中性條件下較易水解[16]。p H 為6.0時,5種物質保留時間差異較大,較易分離。故流動相p H選為6.0。

圖1 流動相p H值對磷酸腺苷和輔酶I保留時間的影響Fig.1 Effects of pH value on the retention time of intracellular adenosine phosphate and coenzyme I
2.2.3 流動相配比選擇考察等度洗脫條件(乙腈體積分數分別為1%、2%、3%、4%和5%)對磷酸腺苷和輔酶I的分離效果,結果見圖2。磷酸腺苷和輔酶I的保留時間隨乙腈比例的增加而減小,乙腈比例較低(<2%)時,NADH難以洗脫出來。當乙腈體積分數為5%時,磷酸腺苷和輔酶I在15 min內被全部洗脫出來,但AMP、NAD+、ADP無法達到良好分離。可見等度洗脫很難滿足分離要求。因此作者選擇梯度洗脫,發現在0~20 min,流動相中三乙胺緩沖液與乙腈的比例由99∶1降低到95∶5,保持5 min,接下來15 min內,流動相比例再由95∶5變化為99∶1時,5種標準物質和樣品均有較好的分離效果,結果見圖3。

圖2 流動相中乙腈體積分數對磷酸腺苷和輔酶I保留時間的影響Fig.2 Effects of different acetonitrile proportion on the retention time of intracellular adenosine phosphate and coenzyme I


圖3 磷酸腺苷和輔酶I標準品的HPLC圖譜(a)和樣品的HPLC圖譜(b)Fig.3 HPLC chromatograms of standards(a)and sample(b)
2.3 工作曲線
分別以各組分峰面積A對標準工作溶液質量濃度c(g/L)進行線性回歸分析,磷酸腺苷和輔酶I的回歸方程、相關系數及檢出限見表2。由表2知,在所測范圍內,線性關系良好。
2.4 精密度和回收率
取一定量待測樣品3份,其中一份做本底,另外兩份分別添加一定量磷酸腺苷和輔酶I的標準混合液,然后按照優化的方法進行檢測。每份樣品進行3次平行測定,計算回收率和精密度,結果表明該方法回收率高,重現性好,見表2。

表2 回歸分析、檢測限、精密度和回收率Tab.2 Regression analysis,detection limit,precision and recovery of the RP-HPLC method
2.5 發酵過程中釀酒酵母胞內磷酸腺苷和輔酶I含量的變化
在上述確定的色譜條件下,檢測發酵過程中下面釀酒酵母和上面釀酒酵母胞內磷酸腺苷和輔酶I的摩爾質量濃度,結果見圖4、5。
ATP普遍存在于生物細胞內,為細胞新陳代謝提供能量,是細胞生存的必要因素。Guimaraes P M R等人用生物發光法研究了厭氧發酵過程中釀酒酵母胞內磷酸腺苷摩爾質量濃度和EC的變化,測得ATP摩爾質量濃度為1.7~4.2μmol/g,ADP摩爾質量濃度為0.1~1.8μmol/g,AMP摩爾質量濃度為0.1~1.8μmol/g,EC 為0.47~0.95[17]。由圖4(A)及圖5(A)可知,本法測定的下面釀酒酵母胞內ATP摩爾質量濃度為1.57~5.32μmol/g,ADP摩爾質量濃度為0.75~1.32μmol/g,AMP摩爾質量濃度為0.54~1.10μmol/g,EC為0.56~0.85。上面釀酒酵母胞內ATP摩爾質量濃度為1.36~5.10μmol/g,ADP摩爾質量濃度為0.36~1.11μmol/g,AMP摩爾質量濃度為0.24~0.88 μmol/g,EC為0.57~0.90,與 Guimaraes P M R測定的結果基本一致。發酵過程中下面釀酒酵母和上面釀酒酵母胞內ATP摩爾質量濃度都是先升高后降低,ADP和AMP摩爾質量濃度都呈升高趨勢。下面釀酒酵母胞內EC在發酵0~3 d內基本不變,而后逐漸降低,而上面釀酒酵母胞內EC在發酵過程中呈逐漸降低趨勢。發酵過程中下面釀酒酵母和上面釀酒酵母胞內ATP、ADP、AMP摩爾質量濃度和EC的變化趨勢都基本一致。由于在發酵開始時,酵母細胞利用麥汁中的營養物質維持代謝,同時合成ATP貯存起來,此時細胞代謝活性較高,ATP摩爾質量濃度較高,EC較高。隨著發酵時間的延長,發酵液中營養物質逐漸被消耗,溶氧量逐漸降低,酵母細胞分解胞內ATP生成ADP、AMP,以供代謝需要,所以胞內ATP摩爾質量濃度逐漸降低,ADP和AMP摩爾質量濃度逐漸升高。此時細胞代謝活性降低,使EC逐漸降低。

圖4 發酵過程中下面釀酒酵母胞內磷酸腺苷(a)及輔酶I(b)摩爾質量濃度的變化Fig.4 Time course of the intracellular adenosine phosphate(a)and coenzyme I(b)during larger Saccharomyces cerevisiae fermentation process
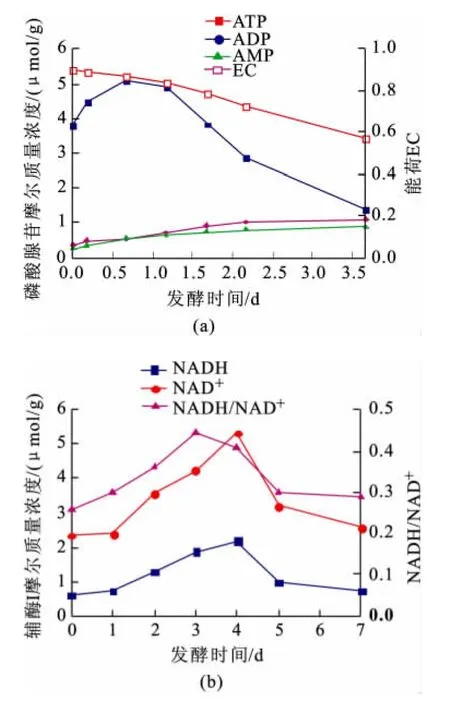
圖5 發酵過程中上面釀酒酵母胞內磷酸腺苷(a)及輔酶I(b)摩爾質量濃度的變化Fig.5 Time course of the the intracellular adenosine phosphate(a)and coenzyme I(b)during ale Saccharomyces cerevisiae fermentation process
NAD+/NADH反映細胞的氧化還原狀態,其水平直接影響著細胞的節律、衰老、癌變和死亡等生命過程[18]。由圖4(b)及圖5(b)可知,發酵過程中下面釀酒酵母和上面釀酒酵母胞內NAD+、NADH摩爾質量濃度和NADH/NAD+都是先升高后降低,下面釀酒酵母和上面釀酒酵母胞內NAD+、NADH摩爾質量濃度和NADH/NAD+的變化趨勢都基本一致。在發酵初期,細胞代謝旺盛,合成較多的輔酶I,細胞還原度較高,但隨著發酵液中的營養物質逐漸被消耗,細胞代謝逐漸減弱,輔酶I摩爾質量濃度逐漸下降,還原度逐漸降低。
3 結語
液氮研磨法能較好地提取胞內代謝物,并能很好地防止ATP和NADH的水解。經過優化,確定0.025 mmol/L pH 6.0的三乙胺-磷酸緩沖液-乙腈作為流動相,梯度洗脫程序為:0~20 min,流動相中三乙胺-磷酸緩沖液與乙腈的比例由99:1降低到95:5,保持5 min,接下來15 min,流動相的比例再由95∶5變化為99∶1。此條件能對胞內磷酸腺苷和輔酶I進行較好地分離,且該方法回收率高,重現性好,可以準確監控發酵過程中酵母胞內磷酸腺苷和輔酶I的代謝情況。結果表明,用液氮研磨法提取,高效液相色譜法測定酵母胞內磷酸腺苷和輔酶I,準確度和精密度均較高,是一種簡便、快速、準確、易于推廣的檢測方法。
(References):
[1]秦耀宗.啤酒工藝學[M].北京:中國輕工業出版社,2008:3.
[2]湖北輕工職業技術學院翻譯組譯.啤酒工藝實用技術(第8版)[M].北京:中國輕工業出版社,2008:297.
[3]劉立明,陳堅,李華鐘,等.氧化磷酸化抑制劑對光滑球擬酵母糖酵解速度的影響[J].生物化學與生物物理進展,2005,32(3):251-257.
LIU Li-ming,CHEN Jian,LI Hua-zhong,et al.Effect of oxidative phosphorylation inhibitors on the glycolytic flux inTorulopsis glabrata[J].Prog Biochem Biophys,2005,32(3):251-257.(in Chinese)
[4]Liu L M,Li Y,Shi Z P,et al.Enhancement of pyruvate productivity inTorulopsis glabrata:Increase of NAD+availability[J].Journal of Biotechnology,2006,126:173-185.
[5]Guimaraes P M R,John L.The adenylate energy charge and specific fermentation rate of brewer's yeasts fermenting high and very high-gravity worts[J].Yeast,2008,25:47.
[6]王翠華,李友元,陳長華,等.溫度對丙酮酸生物合成動力學、能荷和氧化-還原度的影響[J].生物工程學報,2006,22(2):2-6.
WANG Cui-hua,LI You-yuan,CHEN Chang-hua,et al.Effects of temperature on the kinetics and level of energy charge and oxidation-reduction state in pyruvate biosynthesis[J].Chinese Journal of Biotechnology,2006,22(2):2-6.(in Chinese)
[7]王龍耀,范利華,肖安風,等.ATP的分析方法綜述[J].天津化工,2003,17(2):30-33.
WANG Long-yao,FAN Li-hua,XIAO An-feng,et al.Review on the measurement methods of ATP[J].Tianjin Chemical Industy,2003,17(2):30-33.(in Chinese)
[8]Eduardo S,Hugo O,Rita M A,et al.Micromolar HgCl2concentrations transitorily duplicate the ATP level inSaccharomyces cerevisiaecells[J].FEBS letters,2005,579:4044-4048.
[9]張梁,陳蘊,石貴陽,等.HPLC法測定玉米濃醪發酵酒精醪液中的纖維二糖和蜜二糖[J].食品與生物技術學報,2005,24(02):90.
ZHANG Liang,CHEN Yun,SHI Gui-yang,et al.Determination of cellobiose and melibiose in the distillate of high-gravity ethanol broth from corn by HPLC[J].Journal of Food Science and Biotechnology,2005,24(02):90.(in Chinese)
[10]Rasmus D,Britta S,Silas G V,et al.Metabolite profiling for analysis of yeast stress response during very high gravity ethanol fermentations[J].Biotechnology and Bioengineering,2005,6:703-714.(in Chinese)
[11]代瑞華,劉會娟,曲久輝.銅綠微囊藻中磷酸腺苷的提取及分析[J].分析化學,2007,35(12):1701-1705.
DAI Rui-hua,LIU Hui-juan,QU Jiu-hui.Extraction and analysis of adenosine phosphate in cells of microcystis aeruginosa[J].Chinese Journal of Analytical Chemistry,2007,35(12):1701-1705.(in Chinese)
[12]盧群,劉曉艷,丘泰球,等.超聲對酵母細胞膜通透性的影響[J].食品與發酵工業,2005,31(9):14-17.
LU Qun,LU Xiao-yan,QIU Tai-qiu,et al.Effect of ultrasound on membrane permeability ofSaccharomyces cerevisiae[J].Food and Fermentation Industries,2005,31(9):14-17.(in Chinese)
[13]Kazuo S,Yukianri Y,Toshiyuki H,et al.On-line measurement of intracellular ATP ofSaccharomyces cerevisiaeand pyruvate during sake mashing[J].Journal of Bioscience and Bioengineering,2000,3:294-301.
[14]Hugo O,Elisabete C,Mercedes D V,et al.H2O2,but not menadione,provokes a decrease in the ATP and an increase in the inosine levels inSaccharomyces cerevisiae[J].European Journal of Biochemistry,2003,270:1578-1589.
[15]陳永福,王記成,云振宇,等.高效液相色譜法測定傳統發酵乳中的有機酸組成[J].中國乳品工業,2007,35(1):54-58.
CHEN Yong-fu,WANG Ji-cheng,YUN Zhen-yu,et al.Study on organic acids in traditionally fermented milk by HPLC[J].Chinese Dairy Industry,2007,35(1):54-58.(in Chinese)
[16]王鏡巖,朱圣庚,徐長法.生物化學(下冊).第3版[M].北京:高等教育出版社,2007:38.
[17]Guimaraes P M R,Londesborough J.The adenylate energy charge and specific fermentation rate of brewer's yeasts fermenting high-and very high-gravity worts[J].Yeast,2008,25:47-54.
[18]李達,倫永志,周士勝.NAD+/NADH 代謝機制研究進展[J].生物技術通訊,2010,21(1):98.
LI Da,LUN Yong-zhi,ZHOU Shi-sheng.Research progress in NAD+/NADH metabolic mechanisms[J].Letters in Biotechnology,2010,21(1):98.(in Chinese)
Simultaneous Determination of Adenosine Phosphate and Coenzyme I in Cells ofSaccharomyces cerevisiaeby RP-HPLC
LI Hui-pin,ZH AO Mou-ming,YU Zhi-min,LEI Hong-jie,ZH AO Hai-feng*
(College of Light Industry and Food Sciences,South China University of Technology,Guangzhou 510640,China)
In this manuscript,an RP-HPLC method was developed to determine simultaneously adenosine phosphate and coenzyme I in Saccharomyces cerevisiae,in order to monitor metabolic state.The intracellular metabolites extracted by the method of rubbing with liquid nitrogen were separated on an Agilent Zorbax SB-Aq column (5μm,4.6 mm×150 mm).A 0.025 mmol/L triethylamine-phosphoric acid buffer solution(p H 6.0)and acetonitrile mixture was used as mobile phase,with a flow rate of 1 m L/min.Detection wavelength was set at 254 nm.Under these conditions,three kinds of adenosine phosphate and two kinds of coenzyme I could be well separated.The recoveries were between 98.30%and 103.42%,and the RSD of the method was less than 3%.The results showed that this method is sensitive,accurate,reproducible and can be applied to determinate the content of intracellular adenosine phosphate and coenzyme I ofS.cerevisiae.
HPLC,Saccharomyces cerevisiae,adenosine phosphate,coenzyme I
TS 201.2
A
1673-1689(2012)05-0492-07
2011-05-03
國家“十一五”科技支撐計劃項目(2008BAI63B06);廣東省科技計劃工業攻關項目(2009A010700004,2010A010500002)。
*
趙海鋒(1977-),男,河南焦作人,工學博士,講師,主要從事發酵工程與食品生物技術方面的研究。E-mail:hfzhao@scut.edu.cn
