大瀧六線魚(Hexagrammos otakii)熒光AFLP分析體系的建立及優化
2011-12-28 08:17:40仝顏麗李海東王玲王波鄭明剛
海洋通報 2011年6期
仝顏麗,李海東,王玲,王波,鄭明剛
(1.日照職業技術學院,山東 日照 276826;2.青島大學,山東 青島 266061;3.國家海洋局第一海洋研究所 海洋生態中心,山東 青島 266061)
大瀧六線魚(Hexagrammos otakii)熒光AFLP分析體系的建立及優化
仝顏麗1,李海東1,王玲2,王波3,鄭明剛3
(1.日照職業技術學院,山東 日照 276826;2.青島大學,山東 青島 266061;3.國家海洋局第一海洋研究所 海洋生態中心,山東 青島 266061)
以大瀧六線魚為材料,通過對DNA提取質量和濃度、EcoRⅠ/MseⅠ酶切反應時間、Mg2+濃度、dNTPs濃度、引物濃度以及選擇性擴增中預擴增產物稀釋倍數等關鍵因素進行研究,建立并優化了大瀧六線魚熒光AFLP分子標記反應體系。用優化的反應體系,篩選到10對適合大瀧六線魚的引物,為今后利用AFLP標記技術進行大瀧六線魚的遺傳多樣性研究提供了標準化程序。
大瀧六線魚;AFLP;分子標記;反應體系
大瀧六線魚(Hexagraminos otakii)又名歐式六線魚,俗稱黃魚,隸屬于蚰形目(Scorpaeniformes)、六線魚科(Hexagrammidae)、六線魚屬(Hexagrammos)。大瀧六線魚屬冷溫性近海底層魚類,主要分布于我國山東和遼寧等地的近海多巖礁海區以及日本、朝鮮及俄羅斯遠東諸海[1]。六線魚為沿岸淺海定居性魚類,耐低溫、適應能力較強、肉味鮮美,深受國內沿海地區人民的喜愛,已被列為我國名貴海水養殖對象。目前對該魚的研究多集中于其生物學、育苗和營養方面[2-4],遺傳多樣性方面的研究還未見報道。
擴增片斷長度多態性(amplified fragment length polymorphism,AFLP)標記是由荷蘭科學家Zabeau和Vos在PCR和RFLP技術的基礎上發展起來的一種DNA多態性檢測方法,具有RAPD和RFLP技術的雙重優點。AFLP對基因組的多態性檢測不需要預知該基因組的序列特征,易于標準化,被認為是一種理想有效的分子標記技術,現已被廣泛應用于遺傳多樣性分析、品系分析、生態分析等諸多方面[6]。在海洋生物領域,AFLP技術已在櫛孔扇貝[7]、虹鱒[8]、日本對蝦[9]、大黃魚[10]等生物中得到了應用, 但是在大瀧六線魚中的研究還未見報道。
針對不同的試驗材料,建立合適的AFLP反應體系對于試驗結果顯得尤為重要。本研究通過對大瀧六線魚熒光AFLP反應體系進行優化,建立了適合大瀧六線魚的熒光AFLP分析體系,為大瀧六線魚的遺傳多樣性分析及種質資源的保護提供支持。
1 材料與試劑
1.1 實驗材料
本實驗所用大瀧六線魚取自青島近岸水域,實驗樣品液氮速凍后置-80℃冰箱保存。
1.2 試劑與儀器
EcoRⅠ、Mse Ⅰ(NEB),T4 DNA ligase(MBI),Taq酶(TaKaRa),Mse I/EcoR I接頭(上海生工),熒光引物(上海生工),GeneScan-500 LIZ(Applied Biosystems),Hi-Di Formamide(Applied Biosystems),超微量分光光度計(GEHC),PCR擴增儀(Bio-Rad),DL2000 marker(TaKaRa),凝膠成像分析儀(SYNGENE),ABI3130 genetic analyzer(Applied Biosystems),Bullet Blender快速組織細胞破碎儀(Next Advance)
2 方 法
2.1 基因組DNA提取
采用改進的蛋白酶K消化法進行DNA的提取:取-80 ℃冷凍保藏的肌肉組織100 mg,用滅菌的雙蒸水沖洗干凈,用吸水紙將表面的水吸干,置于1.5 mL離心管。加入450 μL 裂解緩沖液(4.5 μL 10 mmol/L Tris-HCl pH=8.0;90 μL 100 mmol/L EDTA pH=8.0和355 μL雙蒸水),于細胞破碎儀將組織破碎,加入10%SDS 150 μL、ProtenaseK 7.5 μL混勻,55 ℃消化(上下緩慢顛倒數次以充分消化至澄清(約3~5 h),加入150 μL NaCl(5mol/L) 混勻,8 000 r/min 離心10 min,吸取上清。加入等體積預冷的異丙醇,混勻,-20 ℃放置30 min,6000 r/min 離心5 min,沉淀用70%乙醇(600~800 μL)洗滌2~3遍。500 μL雙蒸水溶解上述沉淀,加等體積飽和酚液,緩慢來回顛倒混勻10 min,12 000 r/min離心10 min;吸取上層水相,加等體積酚︰氯仿︰異戊醇(25︰24︰1)緩慢顛倒混勻10 min,12 000 r/min離心10 min;吸取上層水相,加等體積氯仿︰異戊醇=24︰1緩慢顛倒混勻10 min,12 000 r/min離心10 min取上清。上清中加入1/10體積2 mol/L NaAc和2倍體積預冷的無水乙醇,-20 ℃放置30 min,12 000 r/min離心10 min去乙醇,500μL 70%乙醇洗滌沉淀,風干加適量雙蒸水溶解。取3 μL DNA進行瓊脂糖凝膠電泳檢測DNA質量,用核酸超微量分光光度計進行定性定量分析。
2.2 酶切體系的優化
酶切反應采用EcoRI和MseI雙酶切組合,酶切體系為:10×NEB buffer 5 μL,100×牛血清白蛋白(BSA)0.5 μL,EcoRI-HF(20 U/μL)1 μL,MseI(10 U/μL)1 μL,模板DNA用量2 μg加雙蒸水至50.0 μL。37 ℃水浴1 h,2 h,4 h,6 h以優化酶切時間,65 ℃水浴20 min,使酶失活。1%瓊脂糖凝膠電泳檢測酶切結果。
2.3 接頭連接體系
酶切產物加入已退火的EcoRI接頭和MseI接頭以及T4 DNA連接酶于16 ℃過夜連接。連接體系如下:酶切產物20.0 μL,T4 Ligase(3 U/μL) 0.2 μL,10×T4 Ligase buffer 0.5 μL,EcoRI接頭(10 μmol/L) 0.5 μL, MseI接頭(100 μmol/L)0.5 μL,加雙蒸水至25.0 μL。
2.4 預擴增反應體系
以連接產物作為模板進行預擴增反應。預擴增反應體系為:10×PCR Buffer 2.0 μL ,dNTPs(各2.5 mmol/L)0.4 μL,E +A(10 mmol/L)0.6 μL,M+C(10 mmol/L)0.6 μL,Taq DNA聚合酶1 U,MgCl(225 mmol/L)1.2 μL,連接產物2 μL,加ddH20至總體積為20 μL 。擴增程序為:94 ℃ 5 min;94 ℃30 s,56 ℃ 1 min,72 ℃ 1 min, 20個循環;72℃10 min。預擴增產物用1%瓊脂糖凝膠電泳檢測。
2.5 選擇性擴增反應體系及其優化
參照預擴增產物的粗略濃度稀釋一定倍數后作為選擴模板,選擴引物為 E+3/M+3引物組合,其中E+3為熒光標記引物。選擴反應體系為:預擴增產物稀釋液5 μL,10×PCR Buffer 2.0 μL, dNTPs(各2.5 mmol/L)1.6 μL,選擇性擴增引物(10 μmol/L)各0.8 μL,Taq DNA聚合酶0.5 U,MgCl2(25 mmol/L)1.6 μL,加ddH20至總體積為20 μL。作者就對選擴結果影響較大的4個因素進行了優化(表1)。 對其中一個因素優化時,保持其它因素不變,比較不同處理結果對擴增結果的影響。
收集2015年1月~2015年12月來我院口腔科門診就診的粘液腺囊腫病人260例,男116例,女144例,年齡16~50歲,囊腫直徑0.5—2.0 cm,其中上下唇105例,舌部155例。所有病例均為首次出現的粘液腺囊腫。

表 1 PCR反應體系中各因素處理方案Tab.1 Different treatments of PCR systems
擴增程序為:94 ℃ 5 min;94 ℃變性30 s,65~56 ℃(每個循環降0.7 ℃)退火30 s,72 ℃延伸1 min,共12個循環;94 ℃變性30 s,56 ℃退火30 s,72 ℃延伸1 min,共20個循環;72 ℃延伸10 min。選擴產物用2%的瓊脂糖凝膠電泳檢測,確定擴增片度大小范圍,用于后續熒光檢測。
2.6 PCR產物的熒光檢測
取選擴產物0.5 μL,加0.25 μL內標(GeneScan TM-500 LIZ1 Size Standard),9.25 μL 去離子甲酰胺(Hi-Di Formamide),混勻后加入96孔板,置于PCR儀95 ℃變性 3 min,立即冰浴5 min。利用POP-7?分離膠在ABI-3130xl自動遺傳分析儀上對擴增結果進行分析。用相應的Run moduLe進行電泳,并接收熒光信號。所得光譜數據用遺傳圖譜4.0軟件(GeneMapper 4.0 software)進行分析。
3 結果與分析
3.1 模板DNA的提取
模板DNA的制備是非常重要的一個環節,若DNA發生斷裂,酶切結果顯然不能代表真實的DNA結構,實驗數據不能反映真實的情況。本實驗采用改進的蛋白酶K消化法進行三次抽提以最大限度的除去雜蛋白,提取結果經過1%瓊脂糖凝膠電泳檢測,條帶清晰明亮(圖1)。經超微量分光光度計測定,各樣品 A260/A280≥1.80,A260/A230≥2.0,表明樣品純度較高,蛋白質等雜質去除較干凈,可以用來進行大瀧六線魚AFLP分析。
3.2 酶切時間對酶切反應的影響
基因組DNA酶切質量是決定AFLP成功與否的關鍵,酶切不充分,所得片段不能涵蓋整個基因組導致所得多態性不真實,因此要對酶切體系進行優化。酶切時間是影響酶切質量的一個重要因素,不同實驗材料因基因組的大小不同,酶切時間長短也有所差異。本試驗用兩個不同的樣品作為模板進行酶切。酶切產物經1%瓊脂糖電泳檢測(圖 2),隨著酶切時間的延長,酶切片段越小濃度越大,酶切6 h所得片度均勻分布在500 bp以下,由于酶切片段在500 bp以下時,在AFLP反應中能被優先擴增,且擴增產物能在凝膠中很好的分離開,便于多態性的檢出。因此本試驗酶切時間選擇6 h。

圖 1 DNA瓊脂糖電泳檢測(1~4道均為大瀧六線魚DNA)Fig.1 Agarose gel electrophoresis of DNA
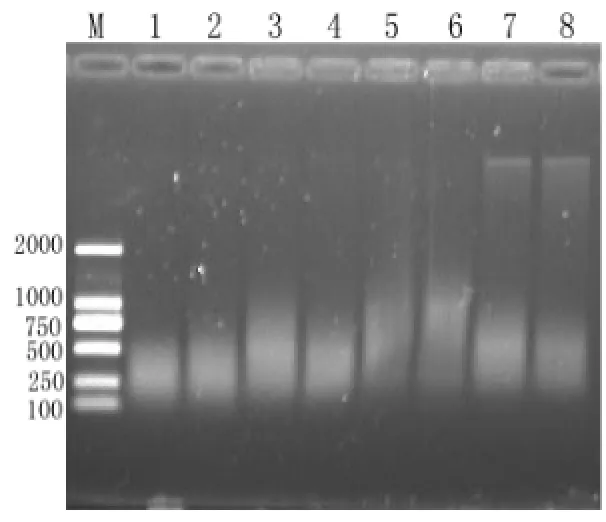
圖 2 基因組雙酶切電泳檢測結果(M:DNA Marker;1,2: 酶切6 h;3,4:酶切4 h;5,6:酶切2 h;7,8:酶切1 h;奇數,偶數分別代表不同樣品的模板)Fig.2 Result of double enzyme digestion at different reaction times
3.3 預擴增體系結果分析
預擴增的質量直接影響選擇性擴增的結果,本實驗設置的反應體系經1%瓊脂糖凝膠電泳檢測(圖 3):其彌散帶主要分布在100~750 bp之間,稀釋后可作為選擇性擴增的模板。
3.4 AFLP選擇性擴增反應體系的優化
3.4.1 預擴增產物的用量對選擇性擴增的影響AFLP對模板濃度要求不高,在濃度相差1000倍的范圍內,得到的結果基本一致,但當模板濃度低于1×10-6μg/μL時得到的AFLP結果往往不可靠[11]。1.0 %瓊脂糖凝膠電泳檢測結果也顯示,一定范圍內,濃度對選擇性擴增的結果影響不大,如圖依次是以稀釋5、10、20、30、50倍預擴產物作為選擇性擴增的模板得到的譜帶,其大小都處在500 bp以下。通過比較,本試驗選擇預擴增產物稀釋30倍作為選擇性擴增的模板。

圖 3 預擴增產物電泳檢測(M:Marker 1~4為不同樣品的預擴增產物)Fig.3 Agarose gel electrophoresis of pre-amplification productions
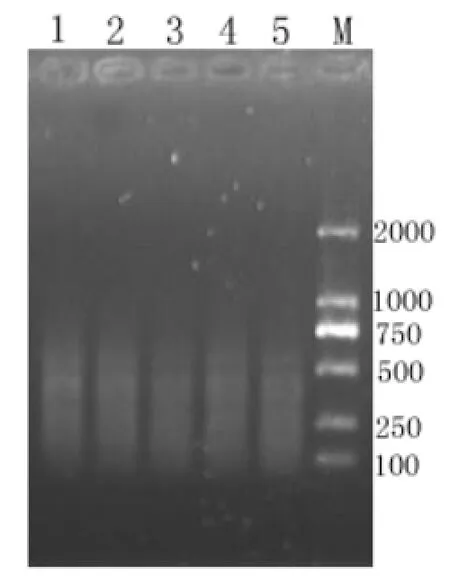
圖 4 不同稀釋倍數預擴增產物的選擇性擴增產物電泳檢測(M: Marker 1~5:分別為稀釋5,10,20,30,50倍的預擴增產物為模板的選擴產物)Fig.4 Detection of the selective amplification products with different diluted templates
3.4.2 dNTPs對選擇性擴增的影響 dNTPs作為擴增產物的原料對擴增效率有密切的影響, dNTPs濃度過高時會使堿基的錯誤摻入率增高,而過低時又會降低PCR產率,本實驗的優化結果表明,dNTPs濃度為0.20 mmol/L時擴增產物清晰且穩定。
3.4.3 引物濃度對選擇性擴增的影響 引物濃度對擴增效率也具有很大的影響,濃度偏高時易出現錯配現象且會增加引物二聚體的合成量,而偏低時擴增出的條帶少,由圖可知:當引物終濃度為0.1~0.2 μmol/L時,擴增條帶少不利于多態性的研究,當引物濃度為0.4~0.5 μmol/L擴增條帶雖然多但非特異性條帶較多影響結果的真實性,而引物濃度為0.3 μmol/L時條帶較多且穩定,因此最佳引物終濃度為0.3 μmol/L。
3.4.4 Taq DNA聚合酶用量對選擇性擴增的影響Taq DNA聚合酶催化PCR產物的合成,其用量過大合成效率較高但易引起錯配而用量過低會降低PCR產物的合成量,因此,優化Taq DNA聚合酶用量對PCR產物的合成量非常重要。當Taq DNA聚合酶用量為0.5 U時,擴出的條帶少,Taq DNA聚合酶用量為1.5~2.5 U時,非特異性條帶較多,而當Taq DNA聚合酶用量為1 U時,結果較好,條帶較多且穩定。所以選擇 Taq DNA聚合酶用量為1 U。
3.4.5 MgCl2濃度對擇性擴增的影響 MgCl2通過影響Taq DNA聚合酶的活性影響選擴結果且對引物和模板的結合及特異性也具有一定的影響[12],MgCl2濃度過高時非特異性條帶較多,MgCl2濃度過小時影響Taq DNA聚合酶的活性擴出的條帶少或無擴增產物,在本擴增體系選擇MgCl2濃度為0.200 mol/L。
3.5 大瀧六線魚AFLP優化反應體系的驗證
本研究為驗證AFLP優化體系的可靠性,以E+AGG/M+CGA為選擇性引物對不同的大瀧六線魚樣品進行選擇性擴增,擴增產物用ABI3130自動遺傳分析儀進行檢測(部分峰譜如圖5所示)。結果表明不同樣品峰譜清晰,且主要集中在100~350 bp范圍內,說明優化的AFLP體系能產生較為理想的結果。
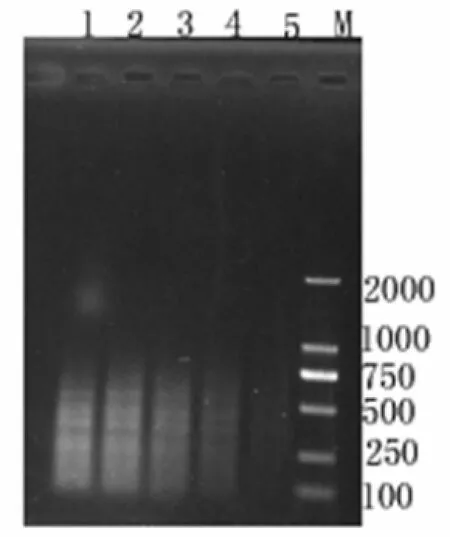
圖 5 不同引物濃度的選擇性擴增結果檢測(M : Marker 1~5分別為引物0.5、0.4、0.3、0.2、0.1μmol/L時的選擇性擴增產物)Fig.5 Detection of the selective amplification products with different primer concentrations

圖 6 不同個體的毛細管電泳檢測結果(1-3代表不同個體在同一選擴引物E+AGG/M+CGA下經基因圖譜分析軟件分析得到的電泳峰譜)Fig.6 Capillary electrophoresis of different individuals
4 討 論
AFLP是4大分子標記(AFLP、SSR、RAPD、RFLP)中公認的最有效的分子標記技術,具有標記多態性強、鑒定靈敏度高、穩定性強、重復性高等諸多優點,是各物種遺傳標記的主要應用技術。
AFLP操作過程較復雜,影響結果的因素較多,不同的材料存在差異,因此筆者針對大瀧六線魚對影響其結果的幾個重要因素進行了優化以期得到真實且準確的結果。高純度的基因組是AFLP成功的最基礎條件,本研究采用改進的蛋白酶K消化法將抽提次數增加到三次,提取的基因組DNA純度較高,各樣品 A260/A280≥1.80,A260/A230≥2.0,符合AFLP技術對基因組的要求。酶切質量直接影響后續擴增的成功與否,50 μL酶切體系中2 μg模板,酶切6 h為宜。預擴反應是對限制性酶切片段的初步篩選即起到對選擴模板純化的作用,預擴產物的質量對后續的選擴影響較大。本試驗就預擴產物的稀釋倍數對選擴結果的影響進行了優化,優化結果顯示:一定范圍內稀釋倍數選擴結果影響不大。對于大瀧六線魚稀釋30倍得到的選擴產物符合預期要求,選擴片度大多處在500 bp以下,亮帶主要集中在100~350 bp。對選擇性擴增體系的優化得到了針對大瀧六線魚的最佳體系即:預擴增產物30倍稀釋液5 μL,10×PCR Buffer 2.0 μL,dNTPs(各2.5 mmol/L)1.6 μL,選擇性擴增引物(10 μmol/L)各0.6 μL,Taq DNA聚合酶1 U,MgCl2(25 mmol/L)1.6 μL,加ddH20至總體積為20 μL。該體系得到的選擴產物穩定且清晰便于觀察。
本研究中,采用F-AFLP(Fluorescent AFLP)分子標記技術,用熒光染料代替傳統的放射性同位素來標記引物,從而得到熒光染料標記的片段,然后用先進的ABI3130遺傳分析儀進行片段多態性的檢測,該方法和放射性同位素及銀染法技術相比,具有高靈敏度、更為快速準確、易于操作且易標準化,可獲得更多的信息,且省時省力效率又高,還可以避免直接用肉眼觀察造成的誤差,其結果也更客觀。
綜上所述,本研究建立了適合大瀧六線魚基因組DNA的AFLP反應體系,為以后利用AFLP技術對大瀧六線魚進行遺傳連鎖圖譜構建、群體遺傳結構及多樣性研究等方面的應用奠定了基礎,以及對其他魚類的相關研究具有借鑒意義。
[1]唐啟升, 葉懋中.山東近海漁業資源開發與保護 [M].北京:農業出版社, 1990.
[2]葉青.青島近海歐氏六線魚食性的研究 [J].海洋湖沼通報, l992, 4: 51-54.
[3]吳立新, 秦克靜, 姜志強, 等.大瀧六線魚人工育苗初步試驗[J].海洋科學, 1996, 4: 32-34.
[4]王書磊, 姜志強, 苗治歐.大連海區大瀧六線魚生物學指標的季節變化 [J].水產科學, 2005, 24(5): 1-3.
[5]Vos P, Hogers R, Bleeker M, et a1.AFLP: a new technique for DNA finger- printing [J].Nucleic Acids Research, 1995, 23(21): 4407-4414.
[6]Vantoai T T, Peng J, Martin S K S.Using AFLP markers to determine the genomic contributions of parents to populations [J].Crop Science, 1997, 37: 1370-1373.
[7]Roa A C, Maya M M, Duque M C, et a1.AFLP analysis of relationships among cassava and other Manihot speices [J].Theoretical and Applied Genetics, 1997, 95: 741-750.
[8]潘潔, 包振民, 趙洋, 等.櫛孔扇貝不同地理群體的遺傳多樣性分析 [J].高技術通訊, 2002, 12: 78-82.
[9]Young W P, Wheller P A, Coryell V H, et al.A detailed linkage map of rainbow trout produced using doubled haploids [J].Genetics, 1998, 148: 839-850.
[10]Moore S S, Whan V, Davis G P, et al.The development and application of genetic markers for the Kuruma prawn Penaeus japonicas [J].AquacuLture, 1999, 173: 19-32.
[11]王志勇, 王藝磊, 林利民, 等.福建官井大黃魚AFLP指紋多態性的研究 [J].中國水產科學, 2002, 9(3): 198-201.
[12]Sambrok J, David W R.分子克隆實驗指南 [M].北京: 科學出版社, 1993.
Establishment and optimization of fluorescence-based AFLP markers forHexagrammos otakiis
TONG Yan-li1, LI Hai-dong1, WANG Ling2, WANG Bo3, ZHENG Ming-gang3
(1.Rizhao Polytechnic, Rizhao 276826, China; 2.Qingdao University, Qingdao 266061, China;3.The First Institute of Oceanography, Qingdao 266061, China)
In this study, we optimized the main factors influencing the results of AFLP, including the quality and concentration of the extracted DNA, reaction time of enzymes EcoRI/MseI digestion, and the concentration of Mg2+, dNTPs, selective primer and the dilute multiple of pre-amplification production were also optimized in selective amplification reaction system.Ten pairs of primers were identified from eighty-one pairs of primers by optimizing the AFLP reaction system.The result established a strong foundation for the AFLP analysis of H.otakii which provided a standardized program for studying genetic diversity of H.Otakii.
Hexagrammos otaki; fluorescence-based AFLP markers; reaction system
P735; S917
A
1001-6932(2011)06-0668-06
2010-08-19;
2011-03-07
國家海洋公益性行業科研專項(201009);國家農業科技成果轉化資金項目(2009-511-208)。
仝顏麗(1989-),女,山東菏澤市人,主要從事海洋生物基因工程研究。電子郵箱:tong111222333@yeah.net。
鄭明剛, 電子郵箱:zmg@fio.org.cn。
猜你喜歡
房地產導刊(2022年5期)2022-06-01 06:20:14
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
建材發展導向(2021年12期)2021-07-22 08:06:48
建材發展導向(2021年7期)2021-07-16 07:07:52
中學生數理化(高中版.高二數學)(2021年12期)2021-04-26 07:43:48
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
海峽科技與產業(2016年3期)2016-05-17 04:32:12
新高考·高一物理(2014年1期)2014-09-18 01:26:07
