副流感病毒感染豚鼠咳嗽反射敏感性變化及其神經源性炎癥機制探討*
2011-10-24 01:47:33葉新民鐘南山劉春麗賴克方陳如沖
中國病理生理雜志 2011年1期
葉新民, 鐘南山, 劉春麗, 賴克方, 陳如沖
(廣州呼吸疾病研究所,廣東 廣州 510120)
·論著·
副流感病毒感染豚鼠咳嗽反射敏感性變化及其神經源性炎癥機制探討*
葉新民#, 鐘南山△, 劉春麗, 賴克方, 陳如沖
(廣州呼吸疾病研究所,廣東 廣州 510120)
目的探討人副流感病毒Ⅲ型(PIV3)感染豚鼠的咳嗽反射敏感性(CRS)變化和機制,以及神經源性炎癥在CRS變化和病毒性咳嗽中的作用。方法豚鼠50只,隨機分成正常對照組以及病毒感染6、12、28和42 d組;通過滴鼻方法接種PIV3。Buxco肺功能儀測定CRS。免疫熒光和FQ-PCR方法檢測PIV3抗原和核酸。ELISA法檢測肺組織勻漿的P物質(SP)含量;FQ-PCR檢測肺組織SP受體NK1、辣椒素受體亞型1(VR1)的mRNA水平;免疫組化方法檢測肺組織SP、VR1和蛋白基因產物(PGP-9.5)的表達及半定量分析,并對它們各自與CRS的相關性進行分析。結果各感染組豚鼠CRS均較對照組明顯升高,以42 d組為最高。免疫熒光可見PIV3抗原表達;FQ-PCR顯示病毒RNA含量在6 d組最高,并隨感染時間逐步降低。各組SP和VR1 mRNA,以及12、28和42 d組NK1 mRNA水平均明顯高于對照組。6、12及28 d組SP,28和42 d組VR1及42 d組PGP-9.5蛋白表達均增強。相關分析顯示,6和28 d組肺勻漿SP含量,12、28和42 d組NK1 mRNA以及各感染組VR1 mRNA均與CRS呈正相關;6、12和42 d組SP的蛋白表達,12和28 d組VR1及12 d組PGP-9.5蛋白表達亦與CRS呈正相關。結論神經肽的釋放增加提示神經源性炎癥的發生。CRS和神經肽的時空變化以及兩者的正相關提示神經源性炎癥在CRS增高和病毒感染性咳嗽中起重要作用。
咳嗽; 咳嗽反射敏感性; 神經源性炎癥; 副流感病毒; P物質
急性上呼吸道感染(upper respiratory tract infection,URI)是急性咳嗽(3周以內)最常見的原因。而感染后咳嗽(URI恢復之后,咳嗽遷延不愈大于3周,而胸片正常)是亞急性咳嗽中的最常見原因[1],一般由病毒感染引起。然而,病毒感染引起的咳嗽,尤其是感染后咳嗽的機制不甚明確,對于治療更是面臨困惑。人類副流感病毒Ⅲ型(parainfluenza virus type 3,PIV3)是人類URI的一個非常常見的病原,絕大部分人在兒童時代已受PIV3感染。血清學監測表示,5歲及以上兒童90%以上有抗人類PIV3抗體。僅僅是因為PIV3感染,就幾乎占美國住院兒童的11%[2]。PIV3感染引起的URI頻繁伴發咳嗽,而且再感染也常常發生。因此對PIV3感染引起咳嗽的機制研究有著重要意義。
咳嗽反射敏感性(cough reflex sensitivity, CRS)異常增高是機體咳嗽反應的一個十分重要的原因。支氣管肺C纖維興奮導致其含有的以P物質(substance P,SP)為主的神經肽釋放引起的神經源性炎癥可能在CRS改變以及咳嗽發病中起重要作用[3]。無髓鞘感覺神經C纖維相當于興奮型的非膽堿能-非腎上腺素能(conadrenergic noncholinergic nerve,NANC)神經。Carr等[4]發現豚鼠副流感病毒感染后表達SP明顯增加。Sekizawa等[5]發現,健康人霧化吸入SP并不直接誘導咳嗽,而呼吸道感染患者吸入SP則可誘發咳嗽。O’Connell等[6]的臨床研究報道,URI患者吸入辣椒素刺激咳嗽后CRS升高。但是,迄今為止,未發現有關URI感染動物的咳嗽和CRS的研究,URI引起CRS增高的機制還不是很清楚。病毒感染可以引起神經肽的異常表達,但這些神經肽在病毒感染性咳嗽發病中的表達和作用究竟如何均有待明確。
辣椒素受體亞型1(vanilloid receptor subtype-1,VR1),是瞬時受體電位通道蛋白的超家族的成員之一。C纖維最顯著特征是纖維末梢上有VR1的功能性表達。VR1受體的活化可誘導C纖維末梢釋放神經肽SP等。有研究發現,慢性咳嗽患者VR1顯著高于健康者;而且,CRS與支氣管黏膜的VR1表達呈高度相關;氣道平滑肌VR1的表達也增強。表明VR1參與了慢性咳嗽的發病過程[7]。但病毒感染后,VR1表達是否增高,以及與CRS的關系如何,目前仍未見報道。有研究發現,呼吸道病毒感染引起蛋白基因產物(protein gene product,PGP-9.5)的表達水平改變,可能是豚鼠肺內發生神經源性炎癥的一個標志[8]。所以本研究也將對PGP-9.5進行檢測。
材 料 和 方 法
1主要實驗試劑及儀器
胎牛血清及MEM、DMEM/F12培養基購自Gibco;辣椒素購自Sigma;P物質ELISA檢測試劑盒購自R&D。RNAisoTMPlus(病毒RNA提取試劑盒)、組織總RNA提取試劑盒、逆轉錄試劑盒及SYBR Green熒光定量PCR反應試劑盒均購自TaKaRa。PIV3 DFI免疫熒光檢測試劑盒購自Millipore。VR1兔抗多克隆抗體、SP和PGP-9.5鼠抗單克隆抗體均購自Abcam。HRP標記免疫組化Ⅱ抗購自Invitrogen;SABC免疫組化試劑盒及DAB顯色試劑盒購自武漢博士德生物有限公司。
單腔非限制型小動物體描儀購自Buxco;MK3型酶標儀購自上海賽默飛世兒儀器公司;Leica DM4000 智能型生物顯微鏡購自Leica。
2細胞培養和PIV3的培養、傳代、凍存及定量
2.1細胞培養 細胞株人恒河猴腎細胞(rhesus monkey kidney epithelial cells,LLC-MK2),來源于ATCC,LLC 細胞是對PIV3敏感的細胞,用于毒株的傳代培養。
2.2病毒的培養、傳代及凍存 PIV3來源于ATCC,LLC細胞在培養瓶中長至70-80%融合后,取病毒懸液200 μL接種于細胞上,每15 min搖動1次,吸附2 h后倒出病毒液,加入2%胎牛血清的培養基,于第2-3 d開始出現細胞病變(CPE),待病變達50-60%時收獲病毒,將病毒反復凍融2-3次后10 000 r/min 4 ℃離心20 min,去除細胞碎片,收集上清分裝后液氮中凍存。
2.3空斑技術測定病毒感染滴度 使用常規空斑技術,培養基為DMEM/F12培養基,以空斑形成單位(plaque-forming unit,PFU)來表示病毒感染滴度。
3方法
3.1實驗動物及分組 雄性純白SPF級豚鼠,購自廣東省實驗動物中心,體重250-300 g之間。隨機分成5組,即正常對照組(vehicle)和4個病毒感染組。每組10只。病毒感染組按病毒感染后天數(post-infection day,PID)又分為6、12、28和42 d 4個組。
3.2動物飼養及病毒接種方法 動物在SPF級環境下飼養,感染組動物單獨飼養,在適應新的環境1周左右后開始接種病毒。病毒滴鼻:乙醚麻醉后,兩側鼻孔分別滴入75 uL的事先擴增凍存好的病毒液。病毒滴度為3.3×105pfu,接種當天同時進行空斑實驗再次鑒定接種病毒的滴度。對照組豚鼠滴入病毒培養用培養基MEM。
3.3豚鼠CRS的測定 本研究應用單腔非限制型小動物體描儀(購自Buxco)來測定豚鼠的CRS,各組動物分別在預定時間進行辣椒素咳嗽激發試驗,判斷CRS的指標主要是咳嗽總次數(cough count,CCnt),參照文獻[9]方法進行操作。Buxco系統通過采集氣流變化信號,計算曲線下面積(V2)以及氣流半數峰值轉換時間,經過模擬方程換算后判斷是否為咳嗽動作,還是打噴嚏或者其它不規則的活動。而辣椒素激發溶液濃度為50 μmol/L,總量為1 mL,觀測記錄10 min(含霧化時間)內的CCnt。
4豚鼠PIV3感染的病毒學檢測
4.1直接免疫熒光法檢測PIV3抗原(操作方法按試劑盒說明書進行) 蘋果綠熒光視為陽性, PIV3主要是胞漿發熒光,可呈不規則的顆粒熒光。
4.2SYBR Green熒光定量PCR(real-time PCR)方法檢測PIV3 RNA的含量(copies/mL) 操作方法按試劑盒說明進行。PIV3上游引物為5’-TTTGCCCTTGGACCGAC-3’,5’- AACCCTGCCCTTTGTGC-3’,產物大小為134 bp;GAPDH作為內參照,上游引物為5’- TGGTGCCAAAAGGGTCA-3’,下游引物為5’- CCTCCACAATGCCGAAGT-3’,產物大小為176 bp。含有和待測樣品相同擴增片段的PIV3病毒質粒DNA標準品由TaKaRa公司合成提供,其病毒核酸含量為2.8×1015copies/L(構建標準品的過程略)。首先提取肺組織病毒RNA,測定濃度和變形瓊脂糖凝膠電泳鑒定質量后,每組取1 μg RNA進行反轉錄反應(RNA逆轉錄為cDNA),20 μL反應體系,反轉錄反應條件為:37 ℃15 min和85 ℃ 5 s。最后進行real-time PCR反應,兩步法PCR擴增標準程序如下:預變性,95 ℃ 30 s 1個循環;PCR反應,95 ℃ 5 s,60 ℃ 30 s,40個循環。結果計算:用已知濃度的病毒標準品10倍稀釋(102-1011copies/L),和未知樣品一起進行PCR反應,通過稀釋標準品得到的標準曲線來確定未知樣品中的病毒拷貝數。
5檢測豚鼠肺組織SP含量的肺組織勻漿液的制備及檢測方法
稱取100 mg新鮮豚鼠肺組織,先加0.1 mol/L醋酸0.3 mL,100 ℃水浴10 min以滅活內源性激肽酶活性,然后按比例加入一定量的PBS,用1-3 mL玻璃勻漿器勻漿,3 500 r/min離心20 min,取上清-70 ℃保存;或水浴后-70 ℃保存,用前勻漿。ELISA法檢測豚鼠肺組織SP的含量(競爭法測抗體):操作及方法按試劑盒說明書進行。
6SYBRGreenreal-timePCR方法檢測豚鼠肺組織NK1、VR1mRNA的相對表達量
NK1上游引物為5’-CTTCCTCCTGCCCTACATCAAC-3’,5’- CTGCCCTGGGTCTGGAAATAC-3’,產物大小為240 bp;VR1上游引物為5’- TCAAAGACCCGGAAACAGGG-3’,5’- CTCTACCAGGAGGGTCACCA-3’,產物大小為224 bp。GAPDH內參照同前,除了先提取組織總RNA和無需標準品做標準曲線外,其余操作方法大致與上文中病毒RNA的檢測方法相同,均按TaKaRa公司的試劑盒說明進行。結果計算:采用GAPDH作為內參照和2-ΔΔCt相對定量法來計算NK1、VR1 mRNA的表達水平。
7免疫組化方法檢測豚鼠肺組織SP、VR1和PGP-9.5蛋白的表達及免疫組化結果圖像分析(半定量方法)
免疫組化圖片用Leica顯微鏡觀察、拍照,進行蛋白表達強度(灰度)的比較。根據以下染色深淺行半定量分析:0=無染色; 1=輕度著色; 2=中度著色;3=強著色;4=很強著色(幾乎呈黑色)。每張片子至少隨機分析5個高倍視野。所有片子均由2個不知情的專業人士分析。
8統計學處理

結 果
1豚鼠CRS測定結果
圖1結果顯示,病毒感染組豚鼠對辣椒素刺激引起咳嗽敏感。感染后6、12、28和42 d組CRS均較正常對照組明顯升高(P<0.05),感染后42 d組較對照組升高最為明顯(P<0.01)。圖2為Buxco系統所顯示的2個相近的典型咳嗽波形;咳嗽過程中的動作和聲音也與人的很相像。


圖1各組豚鼠的CRS比較

Figure 2. The waveforms of two similar typical coughs recorded by the Buxco system.
圖2Buxco儀器記錄的豚鼠2次相近的典型咳嗽波形
2肺組織病理學結果
肺組織外觀無特殊。光鏡下主要是感染組在急性期可見一些氣道炎癥表現,即氣道上皮的腫脹和少許脫落,腺體分泌增加,支氣管壁和肺實質周圍炎癥細胞浸潤,主要是巨噬和淋巴細胞的浸潤增加;而隨著感染后時間延長氣道炎癥逐漸恢復,未見明顯肺實質的炎癥表現(常規病理切片圖省略)。
3PIV3感染的病毒學檢測結果
3.1PIV3在LLC-MK2細胞中培養的CPE作用 PIV3引起細胞病變的典型表現為:感染后2 d左右細胞開始逐漸腫脹、變圓和聚集,最后4-5 d左右直至貼壁細胞破碎、壞死和脫落。
3.2直接免疫熒光技術檢測PIV3病毒抗原結果 與對照組比,PIV3感染的LLC培養細胞可見廣泛的綠色陽性病毒核蛋白抗原的表達;而多數急性感染期6和12d組豚鼠肺組織可見散在綠色的病毒核蛋白抗原的表達,見圖3-6。

Figure 3. The positive results of immunofluorescence in PIV3-infected culture cells of LLC-MK2(×400).
圖3PIV3感染的LLC細胞免疫熒光陽性結果

Figure 4. The negative results of immunofluorescence in normal culture cells of LLC-MK2(×400).
圖4正常培養的LLC細胞免疫熒光陰性結果

Figure 5. The positive results of immunofluorescence in lung tissue section of infected guinea pigs(×200).
圖5感染組豚鼠肺組織免疫熒光陽性結果

Figure 6. The negative results of immunofluorescence in lung tissue section of normal guinea pigs(×200).
圖6正常對照組豚鼠肺組織免疫熒光陰性結果
3.3Real-time PCR方法檢測肺組織PIV3 RNA含量 感染組豚鼠肺組織PIV3 RNA含量在感染后第6 d濃度最高,隨著感染后時間推移,病毒核酸含量呈逐步降低趨勢,很低水平表達維持至感染后42 d(與感染后6 d比,分別P<0.05和P<0.01),見圖7。

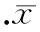
圖7豚鼠肺組織PIV3RNA含量測定結果
4豚鼠肺組織勻漿SP含量的檢測結果
與對照組相比,感染組豚鼠肺組織SP含量感染后6 d起即有明顯升高并達到最高峰(P<0.01),此后直至感染后42 d均維持在高水平(P<0.05),見表1。
5Real-timePCR方法檢測豚鼠肺組織NK1和VR1的mRNA相對表達量結果
PIV3感染后豚鼠肺組織NK1的mRNA水平除了感染后 6 d組豚鼠因為個體差異大而升高沒有統計學意義外,其它感染組其mRNA水平較對照組均有明顯升高(P<0.05),于感染后28 d達到高峰(P<0.01),到感染后42 d仍維持高水平(P<0.01)。VR1的mRNA水平較對照組在感染后均明顯升高(P<0.05),而在28和42 d升高更為突出(P<0.01),見表1。
表1肺組織SP含量及NK1、VR1的mRNA水平

GroupSP(ng/g)▲NK1△VR1△Vehicle19.50±4.681.00±0.471.00±0.34PID631.95±6.35**1.82±1.061.82±0.53*PID1229.32±7.45*2.07±0.93*2.00±0.81**PID2827.91±11.50*2.29±0.94**2.04±0.82**PID4227.12±5.00*2.21±0.74**1.83±0.73*
▲Analyzed by One-way ANOVA and LSD.△Analyzed by Kruskal-Wallis test and Bonferroni test.*P<0.05,**P<0.01vsvehicle group.
6豚鼠肺組織神經肽SP、VR1和PGP-9.5蛋白的免疫組化結果
與對照組(圖8)相比,感染組豚鼠肺組織可見深黃色陽性反應物。表達強的陽性反應顆粒顏色較深,陽性反應細胞數量也顯著多于對照組。按抗體說明書,SP以支氣管上皮、黏膜及分泌物表達為主,見圖9;VR1以細胞質和細胞膜表達,見圖10;PGP-9.5是細胞漿表達,見圖11。免疫組化半定量結果見表2。

Figure 8. A normal result of the immunohistochemistry in animal lung of vehicle group(DAB staining,×400).
圖8完全正常的豚鼠肺組織免疫組化結果

Figure 9. Enhanced expression of SP in PIV3-infected animal(DAB staining,×400).
圖9PIV3感染豚鼠SP強著色

Figure 10. Heavily enhanced expression of VR1 in PIV3-infected animal(DAB staining,×400).
圖10PIV3感染豚鼠VR1很強表達

Figure 11. Heavily enhanced expression of PGP-9.5 in PIV3-infected animal(DAB staining,×400).
圖11PIV3感染豚鼠PGP-9.5很強表達
與對照組相比,感染組SP蛋白陽性表達在感染后6、12及28 d都明顯增強(P<0.05),以28 d組增強最為明顯(P<0.01),而在感染后42 d有所下降,差異沒有統計學意義。感染組VR1蛋白表達在感染后28和42 d組較正常對照組顯著增強(P<0.05)。而感染組PGP-9.5表達在感染后42 d組較對照組明顯增強(P<0.05)。
表2SP、VR1和PGP-9.5蛋白表達的半定量分析
Table 2. The semi-quantitative analysis of SP,VR1 and PGP-9.5 protein expression[(median(interquartile range).n=8)]

GroupSPVR1PGP-9.5Vehicle1.14(0.00)1.14(0.00)1.29(1.50)PID62.25(0.75)*1.71(1.00)2.14(1.50)PID122.25(0.75)*1.67(1.50)2.00(0.50)PID282.38(1.50)**2.14(1.00)*2.43(1.00)PID421.88(0.75)2.20(1.50)*2.80(0.50)*
Analyzed by Kruskal-Wallis and Bonferroni test.*P<0.05,**P<0.01vsvehicle group.
7PIV3感染豚鼠CRS與神經肽相關性分析
7.1CRS與肺組織SP蛋白含量和NK1及VR1 mRNA水平的相關性分析 對于肺組織勻漿SP蛋白含量,病毒感染6和28 d組與CRS呈正相關;對于NK1 mRNA表達,病毒感染12、28和42 d組與CRS呈正相關,其中42 d組相關程度最高(rs=0.8810,P<0.01);對于VR1 mRNA,豚鼠CRS均與之呈正相關(P<0.05),見表3。
7.2CRS與肺組織SP、VR1和PGP-9.5蛋白表達的相關性分析 對于肺組織SP蛋白表達,6、12和42 d組與CRS呈正相關(P<0.05);對于VR1,12和28 d組與CRS呈正相關(P<0.05);而對于PGP-9.5,12 d組與CRS密切正相關(P<0.05)。因為每組只有7例較理想的免疫組化切片可以進行相關比較,所以n=7,見表4。
表3CRS與SP含量和NK1、VR1mRNA水平的相關性分析
Table 3. The correlation analysis between CRS and SP content or NK1,VR1 mRNA levels(n=8)

GroupSPNK1VR1PID6rs0.7110.6670.783P<0.05>0.05<0.05PID12rs0.3810.7380.786P>0.05<0.05<0.05PID28rs0.8050.7560.805P<0.05<0.05<0.05PID42rs0.6670.8810.810P>0.05<0.01<0.05
表4CRS與SP、VR1和PGP-9.5蛋白表達的相關性分析
Table 4. The correlation analysis between CRS and the protein expression of SP,VR1 and PGP-9.5(n=7)

GroupSPVR1PGP-9.5PID6rs0.7650.3420.609P<0.05>0.05>0.05PID12rs0.7090.7370.866P<0.05<0.05<0.01PID28rs0.6710.7590.613P>0.05<0.05>0.05PID42rs0.7840.5750.016P<0.05>0.05>0.05
討 論
病毒感染使興奮型非腎上腺素能非膽堿能(exitatory NANC,eNANC)神經的作用增強,釋放神經遞質,即各種感覺神經肽增多,可使咳嗽中樞的興奮性增高,可使機體對以前并不產生反應的一些刺激敏感性增高等,可能是病毒感染引起CRS增高的咳嗽的外周和中樞性機制。雖然目前仍缺乏直接證據,但有研究在PIV3引起URI的過程中,對神經肽水平、速激肽受體及它們功能的影響方面,發現感染豚鼠氣道主要是在感染后對Ach和SP呈現高反應性,SP的釋放和氣道高反應性(airway hyperreactivity,AHR)一致[10]。Carr等[4]的研究中發現,SP從感染急性期表達開始逐漸增加,而在感染后第28 d到達高峰,當病毒引起的氣道炎癥幾乎已經消失的時候,SP的表達隨后才逐漸降低。由此可以證明病毒感染后,eNANC神經釋放感覺神經肽增多。病毒感染引起的eNANC神經功能增強不僅對AHR,對引起咳嗽尤其是感染后咳嗽的機制也是重要的。釋放的神經肽可以產生一系列外周氣道炎癥表現,產生的神經源性炎癥刺激呼吸道咳嗽感覺神經末梢,導致咳嗽發生,就可能在病毒性咳嗽中發揮重要作用。病毒感染使感覺神經肽增高的原因則可能一方面是因氣道上皮損傷使SP等易于透過黏膜上皮或直接刺激引起分泌增加;同時,由于上皮損傷,上皮細胞合成中性內肽酶減少,使得神經肽降解減少,感覺神經肽堆積而作用增強[11]。
Jacoby等[12]的研究發現,PIV導致的M2受體失活和AHR的作用,可以用NK1受體拮抗劑拮抗而用具有同樣抗炎作用的NK2受體拮抗劑則無效,說明PIV感染主要是釋放的SP及其受體NK1在其致炎過程中起作用。本文雖然沒有進行M2受體方面的研究,但對SP及其NK1受體的研究發現,PIV3感染后,肺勻漿SP蛋白含量在第6 d即明顯增高并達到高峰,直到42 d仍很高;除了感染后第6 d外,NK1 mRNA的表達水平逐步明顯上調,在感染后28 d達到最強。因此說明SP及其NK1受體在PIV3感染引起的咳嗽,尤其是感染后咳嗽中起重要作用。雖然SP蛋白表達在42 d組,可能是因為組織學半定量不那么準確,顯示較對照組無明顯增強外,其它時點均與SP的含量相類似。與CRS的關系研究結果顯示,SP含量的升高和蛋白表達的增強與CRS的升高大多呈正相關,而NK1 mRNA水平與CRS的變化多呈正相關。因此本研究表明,SP及其NK1受體的釋放增加可能是引起PIV3感染性咳嗽CRS增高的主要機制之一。
Watanabe等[13]研究發現免疫組織化學結果顯示VR1常與神經肽在同一軸突中共存在,豚鼠氣管上皮內的VR1大部分與SP共分布。一些研究證實VR1與咳嗽直接相關,氣管上皮內的軸突末梢及上皮下平滑肌和血管,甚至細支氣管和肺泡周圍均見VR1表達[14]。氣道炎癥引起的組織充血、水腫,細胞變性、壞死等改變可在局部形成一個含質子的低pH和中熱的環境,使咳嗽受體的興奮性增加,合成和釋放更多的VR1。多種炎性介質也能直接致敏VR1受體,形成惡性循環,因此氣道炎癥最終可能使咳嗽感受器發生重塑性改變。同Watanabe的研究類似,本文的研究不但發現VR1的蛋白表達在感染后28和42 d明顯增強;而基因表達結果顯示都是在感染后明顯上調;感染各組VR1 mRNA的水平全部與CRS呈正相關。因此本研究證明了VR1可能也是PIV3感染引起CRS增加和神經源性炎癥的一個重要原因。
目前,有關感染后咳嗽的發病機制仍然知之甚少,一般認為是由于病毒感染引起上氣道和/或下氣道的廣泛炎癥導致上皮完整性受到破壞,伴或不伴AHR。有對于呼吸道合胞病毒的研究證明,低劑量(105-106PFU)感染主要是引起支氣管炎,而沒有肺炎發生[15]。參考這個標準,我們選擇了3.3×108PFU/L 的PIV3劑量150 μL進行了預實驗,發現這一劑量能引起豚鼠CRS明顯增加,而病理顯示沒有引起肺炎,因此確定這個劑量可以作為豚鼠病毒感染咳嗽的劑量。我們主要目的是導致豚鼠輕度感染,只引起URI或者最多伴發支氣管炎而盡量不引起肺炎。肺組織病理學結果證明,PIV3感染急性期表現為氣道炎癥改變為主,因此存在一過性或者輕度下氣道的炎癥,與感染后咳嗽的診斷是符合的。但關鍵問題是,通常一般的病毒性URI引起的氣道上皮炎癥在逐步恢復,28 d時已經不很明顯了,而神經肽方面的研究發現,它們的改變顯示特定的時空分布特點,引起的神經肽水平升高多在28 d達到高峰,直到42 d仍很明顯;而且與CRS的變化也基本一致,說明感染引起的神經源性炎癥是CRS升高和感染后咳嗽的關鍵,因此也解釋了PIV3為什么容易引起較長期咳嗽的原因。
有關代表呼吸道總神經纖維分布,包括支配SP等神經肽的神經纖維分布的PGP-9.5,在咳嗽,尤其是病毒感染引起的咳嗽中的作用研究目前仍無報道。本研究中發現PIV3感染后,豚鼠肺組織PGP-9.5中明顯表達增強。雖然它們的蛋白表達與CRS的增高多不相關,因此需要更多的研究,但足以表明PIV3感染后存在氣道神經纖維分布和構成的改變甚至重塑,其結果就會導致與咳嗽相關的神經功能的改變,主要是eNANC神經的功能增強,產生了以SP為主的神經肽引起的神經源性炎癥,可能是病毒性咳嗽的關鍵機制。病毒感染最終一方面通過引起的外周氣道炎癥直接導致咳嗽;另一方面通過增加神經肽表達,引起神經源性炎癥使外周咳嗽感受器敏感化。增加的神經肽使神經末梢的感覺敏感性增強及向腦干輸入沖動增強引起咳嗽閾值下調等直接和間接的作用,在CRS增高或咳嗽反射可塑性變化中發揮重要作用。
綜上所述,本研究發現,PIV3感染引起豚鼠對辣椒素刺激敏感,CRS明顯增高。SP含量增高和蛋白表達增強,NK1 mRNA和VR1 mRNA水平升高,以及VR1和PGP-9.5的表達增強均提示感染引起了神經源性炎癥。多數感染組SP、NK1和VR1與CRS緊密相關,可能是引起后者的關鍵因素。表明在PIV3感染引起的咳嗽中,氣道神經源性炎癥在其發病機制中起決定作用。
[1] Kwon NH, Oh MJ, Min TH, et al. Causes and clinical features of subacute cough[J]. Chest, 2006, 129(5):1142-1147.
[2] Moscona A. Entry of parainfluenza virus into cells as a target for interrupting childhood respiratory disease[J]. J Clin Invest, 2005, 115(7):1688-1698.
[3] Lee LY. Respiratory sensations evoked by activation of bronchopulmonary C-fibers[J]. Respir Physiol Neurobiol, 2009, 167(1):26-35.
[4] Carr MJ, Hunter DD, Jacoby DB, et al. Expression of tachykinins in nonnociceptive vagal afferent neurons during respiratory viral infection in guinea pigs[J]. Am J Respir Crit Care Med, 2002, 165(8):1071-1075.
[5] Sekizawa K, Jia YX, Ebihara T, et al. Role of substance P in cough[J]. Pulm Pharmacol, 1996, 9(5-6):323-328.
[6] O’Connell F, Thomas VE, Studham JM, et al. Capsaicin cough sensitivity increases during upper respiratory infection[J]. Respir Med, 1996, 90(5):279-286.
[7] Groneberg DA, Niimi A, Dinh QT, et al. Increased expression of transient receptor potential vanilloid-1 in airway nerves of chronic cough[J]. Am J Respir Crit Care Med, 2004,170(12):1276-1280.
[8] Tan Y, Yang T, Liu S, et al. Infection with respiratory syncytial virus alters peptidergic innervation in the lower airways of guinea-pigs[J]. Exp Physiol, 2008, 93(12):1284-1291.
[9] Lewis CA, Ambrose C, Banner K, et al. Animal models of cough: literature review and presentation of a novel cigarette smoke-enhanced cough model in the guinea-pig[J]. Pulm Pharmacol Ther, 2007, 20(4):325-333.
[10]Kudlacz EM, Shatzer SA, Farrell AM, et al. Parainfluenza virus type 3 induced alterations in tachykinin NK1 receptors, substance P levels and respiratory functions in guinea pig airways[J]. Eur J Pharmacol, 1994, 270(4):291-300.
[11]Jacoby DB. Pathophysiology of airway viral infections[J]. Pulm Pharmacol Ther, 2004, 17(6):333-336.
[12]Jacoby DB, Yost BL, Elwood T, et al. Effects of neurokinin receptor antagonists in virus-infected airways[J]. Am J Physiol Lung Cell Mol Physiol, 2000, 279(1):L59-L65.
[13]Watanabe N, Horie S, Michael GJ, et al. Immunohistochemical co-localization of transient receptor potential vanilloid (TRPV)1 and sensory neuropeptides in the guinea-pig respiratory system[J]. Neuroscience, 2006, 141(3):1533-1543.
[14]Morice AH, Geppetti P. Cough· 5: The type 1 vanilloid receptor: a sensory receptor for cough[J]. Thorax, 2004, 59(3):257-258.
[15]Dakhama A, Lee YM, Gelfand EW. Virus-induced airway dysfunction: pathogenesis and biomechanisms[J]. Pediatr Infect Dis J, 2005, 24(11 Suppl):S159-S169.
Thechangesofcoughreflexsensitivityinguineapigswithparainfluenzavirusinfectionanditsneurogenicmechanism
YE Xin-min, ZHONG Nan-shan, LIU Chun-li, LAI Ke-fang, CHEN Ru-chong
(GuangzhouInstituteofRespiratoryDiseases,Guangzhou510120,China.E-mail:nanshan@vip.163.com)
AIM: To investigate the changes of cough reflex sensitivity (CRS) in guinea pigs of human parainfluenza virus type 3(PIV3) infection and the mechanisms, as well as the role of neurogenic inflammation in CRS change and the pathogenesis of viral cough.METHODSFifty guinea pigs were randomly divided into normal control group and four virus infection groups of post-infection day (PID) 6,12,28 and 42. PIV3 was inoculated by intranasal instillation. The Buxco system was used to assess cough reflex sensitivity (CRS). PIV3 antigen and nucleic acid were detected by immunofluorescence and fluorescence quantitative PCR (FQ-PCR). Substance P (SP) content in lung homogenates was assayed by ELISA. NK1 receptor of SP and vanilloid receptor subtype 1 (VR1) mRNA levels in lung tissue were determined by FQ-PCR. The protein expression of lung SP, VR1 and protein gene product (PGP-9.5) were examined by immunohistochemistry and semi-quantitative analysis. The correlation between neuropeptides and CRS was analysed.RESULTSThe CRS of infected guinea pigs was significantly higher than that in control group, with the highest in PID 42. The viral antigen expression can be seen in both PIV3-infected culture cells and lung tissues. The content of PIV3 RNA peaked in PID 6 and showed a decreasing trend with infection time. The SP and VR1 mRNA in all infected groups and NK1 mRNA levels in groups of PID 12,28 and 42 increased significantly. The protein expression of SP in PID 6,12 and 28, VR1 in PID 28 and 42 and PGP-9.5 in PID 42 enhanced obviously. Correlation analysis showed that the content of SP in PID 6 and 28, NK1 mRNA in PID 12, 28 and 42 and VR1 mRNA in all infected groups had a positive correlation with CRS.The protein expression of SP in PID 6,12 and 42, VR1 in PID 12 and PGP9 in PID 12 showed a positive correlation with CRS.CONCLUSIONThe increase of neuropeptides releasing in PIV3-infected groups suggested that neurogenic inflammation was caused. The temporal and spatial variation of CRS and neuropeptides and the positive correlation between both indicated that airway neurogenic inflammation may play a decisive role in the increase of CRS and postinfectious cough caused by viral infection.
Cough; Cough reflex sensitivity;Neurogenic inflammation;Parainfluenza virus; Substance P
R56
A
10.3969/j.issn.1000-4718.2011.01.001
1000-4718(2011)01-0001-08
2010-04-23
2010-06-11
國家自然科學基金資助項目(No.30700341)
△通訊作者 Tel:020-83328322; E-mail: nanshan@vip.163.com
#現工作單位:南京兒童醫院呼吸科
