多糖激活巨噬細胞的信號轉導通路
2011-10-22 10:28:25馬志新趙魯杭綜述
浙江大學學報(醫學版) 2011年5期
關鍵詞:信號
馬志新,趙魯杭 綜述
多糖激活巨噬細胞的信號轉導通路
馬志新,趙魯杭 綜述
(浙江大學醫學院生物化學與遺傳學系,浙江杭州310058)
多糖是一類來源廣泛,通過多途徑多機制激活巨噬細胞而發揮免疫調節功能的天然活性物質。闡明來源不同(植物、真菌、藻類)的多糖激活巨噬細胞的信號傳導通路,不僅有助于從分子水平了解其調節免疫功能的作用機制,也可為新型靶向免疫調節藥物的開發提供新的方向。
多糖類/藥理學;巨噬細胞/免疫學;信號傳導
[JZhejiang Univ(Medical Sci),2011,40(5):567-572.]
多糖是由單糖聚合成的線性或分支的聚合物,具有顯著的免疫調節、抗腫瘤、抗炎癥、抗病毒、抗輻射等多種生物學功能,已被應用于消化道黏膜損傷和潰瘍、血液系統疾病、組織挫傷及燒傷等多種疾病的治療中[1]。由于多糖具有顯著的免疫調節功能臨床應用廣泛,而且相對低毒,已經成為新型免疫調節劑和基因輔助細胞治療藥物開發的重點領域[2]。巨噬細胞在真核生物種系間高度保守,發揮抗原遞呈及協同T淋巴細胞調節適應性免疫反應[3];它通過免疫監視、趨化作用、吞噬作用及對靶細胞的直接殺傷參與免疫調節,而且其與胚胎發生過程中的組織塑形、損傷修復和衰老細胞的清除有關[4]。研究證實,不同多糖由于其糖鏈結構(α1,6、α1,4、β1,4)和組成(單糖種類和比例)的差異對巨噬細胞的激活效應也不同。多糖以半乳糖、鼠李糖和阿拉伯糖為主要成分,其分子量大于100 kD的多糖相對于低分子量多糖具有較高的生物活性,能顯著促進巨噬細胞對一氧化氮(NO)和多種細胞因子的生成,以增強宿主的免疫力[5-6]。
多糖激活巨噬細胞的方式主要通過2條途徑。第一,受體介導的激活作用:多糖經受體的介導,并經通路下游分子將活化信號傳入巨噬細胞,啟動免疫應答。巨噬細胞表面與多糖的識別相關受體主要有 Toll樣受體(TLRs)、CD14、補體3受體(CR3)、清道夫受體(SR)、甘露糖受體(MR)、Dectin-1和G蛋白偶聯受體。第二,內吞作用介導的巨噬細胞激活。前者在巨噬細胞的激活中居主導地位,后者通常發揮協同作用。
1 多糖激活巨噬細胞的信號轉導通路
多糖通過模式受體(CR3、TLRs、Dectin-1、MR、SR)、CD14及不同受體間通過形成信號轉導復合物(TLR4-CD14、CR3-CD14)相互協作激活巨噬細胞的數條信號通路(圖1)。
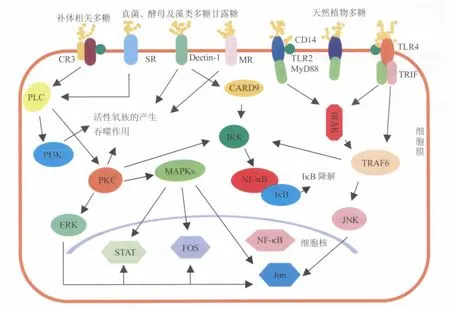
圖1 受體介導的多糖激活巨噬細胞的信號通路Fig.1 Signaling pathway of polysaccharide on macrophage mediated by receptors
1.1 TLR4介導的信號轉導通路 TLRs是一類新型跨膜蛋白樣模式識別受體,廣泛表達于樹突狀細胞、巨噬細胞,中性粒細胞和淋巴細胞的表面,是目前研究免疫功能調節的熱點[7]。TLR4作為TLRs家族的重要成員,是多糖、生物堿、植物黑色素等多種天然活性成分的識別位點[8],TLR4與配體結合后,在髓樣分化蛋白(MyD88)的介導下啟動MyD88依賴的信號傳導通路,促進白細胞介素-1(IL-1)受體相關激酶(IRAK)發生自身磷酸化,激活腫瘤壞死因子受體相關分子-6(TRAF-6),使有絲分裂原激活酶家族(MAPKs包括 ErK1/2、JNK、P38)活化,從而激活核轉錄因子-kappaB(NF-κB)誘導激酶(NIK),NIK 再激活 NF-κB 抑制蛋白(IκB)激酶(IKK),使 IκB 降解并釋放出 NF-κB,啟動或增強靶基因的轉錄,提高巨噬細胞免疫功能[9]。
紅花多糖、刺五加多糖、豬苓多糖、桔梗多糖和云芝多糖都是經該受體激活巨噬細胞。上述多糖作用于TLR4野生型和TLR4缺陷型巨噬細胞的研究發現,后者對多糖的刺激呈低反應性,免疫活性分子的生成明顯受阻,這可能與TLR4表達的缺失密切相關[10-14]。
1.2 CD14和CR3介導的信號轉導通路CD14對脂多糖(LPS)有高度親和力;CR3是由CD11b和CD18兩個亞基組成的黏附分子,能識別β-葡聚糖。在巨噬細胞活化的信號傳導通路中CD14和CR3通常與其他多種受體(例如Dectin-1、TLRs)形成信號傳導復合體,跨膜傳遞活化信號[15]。CD14和 CR3與多糖識別結合后激活磷脂酶C(PLC),產生蛋白激酶C(PKC)和磷脂酰肌醇3-激酶(PI3K),使MAPKs發生磷酸化,進而活化信號傳導及轉錄激活因子(STAT)和NF-κB,上調多種細胞因子的基因表達和釋放[16]。
茯苓多糖與巨噬細胞表面CD14及CR3相結合后,通過上調誘導型一氧化氮合酶(iNOS)的表達,增強巨噬細胞的免疫活性;而CD14及CR3特異性抗體可以使該激活效應降低39%。這說明除CD14和CR3外,茯苓多糖還通過其它途徑活化巨噬細胞[17]。非水溶性的大分子酵母β-葡聚糖(WGP)能使巨噬細胞內酪氨酸激酶(Syk)磷酸化水平顯著升高,促進多種免疫活性分子的釋放。進一步研究發現,WGP可以增加CR3基因缺陷型巨噬細胞對細胞因子的釋放,而MyD88基因缺陷型巨噬細胞對其卻呈現低反應性。所以CR3不是WGP的重要模式識別受體,而有可能是通過MyD88依賴性通路或者Syk途徑來激活巨噬細胞。這雖然與傳統認為的CR3是β-葡聚糖受體相悖,但是這為研究非水溶性大分子多糖激活巨噬細胞的機制提供了新的方向[18]。
1.3 SR介導的信號轉導通路 SR由2個跨膜結構域、2個膜內結構域和1個膜外結構域構成;其與巨噬細胞吞噬清除功能及分泌細胞因子相關,是褐藻多糖硫酸酯、墨角藻聚糖發揮抗炎作用的有效靶點。目前研究顯示,SR可能與CR3通過相同的通路激活巨噬細胞[19-20]。
Nakamura首次發現,墨角藻聚糖能刺激野生型(SR+/+)巨噬細胞釋放NO,而對SR基因缺陷型(SR-/-)巨噬細胞則無誘導作用。經SR活化的巨噬細胞通過P38和NF-κB,這2條相互獨立的信號通路誘導巨噬細胞iNOS活性增強,促進NO釋放。通過阻斷SR膜內結構域可以完全抑制墨角藻聚糖誘導的巨噬細胞P38磷酸化,阻斷NF-κB與DNA的結合,使細胞因子相關基因的表達受限,這說明墨角藻聚糖只通過SR一種受體與巨噬細胞結合[21]。隨后,Jin等證實墨角藻聚糖通過SR可以選擇性地抑制蛋白活化因子-1的激活,下調LPS誘導的iNOS基因表達,起到阻斷NO釋放和抗炎作用。這說明SR在啟動免疫應答的過程中也參與了免疫負反饋調節,避免免疫反應過強造成的細胞毒性反應[22]。
1.4 Dectin-1介導的信號轉導通路 Dectin-1由4亞基組成,是酵母多糖、真菌多糖及β-葡聚糖的模式識別受體。其通常與TLRs協同激活巨噬細胞,經多條通路啟動巨噬細胞的吞噬作用、活性氧族(ROS)的產生及細胞因子的合成釋放[23-25]。Dectin-1與配體結合后其胞漿側免疫受體酪氨酸激活基序(ITAM)激活Syk,Syk促進天冬氨酸特異性的半胱氨酸蛋白水解酶-9(CARD9)結構域發生變化,進而調節與CARD9結合的IKK磷酸酶激活復合體,通過IKK 降解 IκB,釋放 NF-κB,啟動對巨噬細胞的活化作用[26]。靈芝孢子多糖(GSG)對Dectin-1通路缺陷型巨噬細胞激活能力遠低于其對野生型的作用,GSG還通過 TLRs使 MAPKs磷酸化,促進細胞因子的釋放[27]。蟲草多糖(SCG)誘導的細胞因子分泌在Dectin-1敲除巨噬細胞中被完全阻斷,但卻不受MyD88缺陷的影響。這表明與GSG同時啟動Dectin-1和TLRs不同,SCG對巨噬細胞激活是單一依賴Dectin-1的,這可能是由于SCG與GSG組成成分差異所致[28]。
1.5 MR介導的信號轉導通路 MR是由5亞基組成的C型凝集素,胞外結構域能夠識別甘露糖、L-巖藻糖及N-乙酰葡萄糖氨,在TLR2的協同下參與巨噬細胞的抗原遞呈,啟動免疫應答,目前關于MR通路下游的分子機制還不明確[29]。
蘆薈多糖含有91.5%的甘露糖,經MR激活巨噬細胞,引起巨噬細胞表面主要組織相容性抗原Ⅱ類分子(MHC-Ⅱ)和 IgGFc段受體(FcγR)的表達上調,MHC-Ⅱ可以提高巨噬細胞的抗原呈遞能力,FcγR則增強巨噬細胞清除抗原抗體復合物或者抗體腫瘤細胞復合物[30]。
1.6 G蛋白偶聯受體介導的信號轉導通路多糖與G蛋白偶聯受體胞外結構域識別結合后,受體胞內結構域通過與G蛋白偶聯調節相關酶活性,在胞內產生第二信使,從而將胞外信號跨膜轉入胞內。G蛋白偶聯受體所介導的細胞信號轉導包括cAMP信號通路和磷脂酰肌醇信號通路。
靈芝多糖(GLB7)能引起小鼠腹腔巨噬細胞中cAMP濃度快速升高,蛋白激酶A(PKA)活性也明顯上升[31]。這表明GLB7與G蛋白偶聯受體結合后,激活胞內腺苷酸環化酶(AC)催化ATP脫去一個焦磷酸生成cAMP,而cAMP作為第二信使激活PKA,促進細胞DNA的合成,巨噬細胞的分裂增殖。此外,靈芝多糖還能引起小鼠腹腔巨噬細胞內三磷酸肌醇(IP3)和二酰甘油(DG)濃度的升高,證明IP3/Ca2+和DG/PKC兩條通路均參與了靈芝多糖對巨噬細胞免疫功能的調節[32]。
1.7 多糖激活巨噬細胞的其它通路 除了受體介導的激活通路外,多糖還可以通過巨噬細胞的吞噬作用直接入胞,被吞噬的多糖分子不易完全酶解進而保留其免疫原性,可以協同受體介導的信號通路共同激活巨噬細胞。此外,有報道證實,部分多糖還可以通過TLR6、TLR2及糖皮質激素誘導的腫瘤壞死因子受體(GITR)激活巨噬細胞[33-34]。
SR和CR3通過活化通路下游信號分子PLC、PKC 及 PI3K 激活 MAPKs、STAT、NF-κB,最終觸發相關基因轉錄的起始。Dectin-1、MR信號通路的激活可以促進巨噬細胞吞噬作用、ROS的產生和NF-κB的釋放。TLRs在MyD88的介導下激活 IRAK,進而活化 TRAF-6和MAPKs,最終激活 NF-κB。JNK主要存在于胞質內,在細胞受到刺激后,JNK迅速聚積于核內,并導致相應基因表達改變,FOS是一類參與轉錄的調節蛋白。多糖經上述通路激活炎性細胞因子前體和iNOS基因的表達,調節巨噬細胞的免疫功能。
2 多糖對巨噬細胞功能的影響
激活的巨噬細胞的吞噬能力和對外源性物質的清除殺傷能力均顯著增強,對腫瘤細胞的細胞毒性作用明顯提高,細胞因子的分泌和細胞表面相關模式受體的表達上調,總體上提高其免疫應答能力。例如,人參多糖不僅能促進腹腔巨噬細胞產生 ROS、NO、TNF-α 和 IL-6,還能明顯增強其對中性粒細胞的吞噬及細胞表面CD68的表達,提高對K562、HL-60及KC1α腫瘤細胞的細胞毒性作用[35]。酵母多糖激活肺泡巨噬細胞,產生 TNF-α和 IL-10,并且上調TLR4、TLR2 及 Dectin-1 的表達[36]。近年來多糖衍生物對免疫功能調節的研究也受到眾多學者的關注,多糖經化學修飾后對巨噬細胞的激活作用顯著增強,但是,衍生所得多糖的生物安全性還需進一步驗證[37]。
綜上所述,多糖對巨噬細胞免疫功能調節的高效性和低毒性使其在免疫藥物開發領域備受關注。多糖活化巨噬細胞的通路十分復雜,而且不同受體介導的通路之間往往相互調控。從細胞整體水平上來看,一種多糖可能同時與多種受體結合,激活數條信號傳導通路,同時又由于多糖與受體的親和力的差異,導致不同傳導通路對巨噬細胞激活效應的不同。目前對于多糖的研究還處于起始階段,由于其分子自身結構的復雜性和多樣性,對多糖結構測定,表征鑒定都提出了巨大挑戰,多糖分子不同結構域及糖鏈中不同手性的碳原子與受體結合的特異性還鮮為了解。進一步深入了解多糖分子的組成和結構,并闡明其激活巨噬細胞信號通路,對于多糖類靶向免疫調劑的開發有指導意義。
[1] LEUNG M Y,LIU C,KOON J C,et al.Polysaccharide biological response modifiers[J].Immunol Lett,2006,105(2):101-114.
[2] DONG L,XIA SH,LUO Y,et al.Targeting delivery oligonucleotide into macrophages by cationic polysaccharide from bletilla striata successfully inhibited the expression of TNF-alpha[J].Journal of Controlled Release,2009,134(3):214-220.
[3] BIRK R W,GRATCHEV A,HAKIY N.et al.Alternative activation of antigen-presenting cells:concepts and clinical relevance [J].Hautarzt,2001,52(3):193-200.
[4] BEUTLER B.Innate immunity:an overview [J].Molecular Immunology,2004,40(12):845-859.
[5] LO T C T,JIANG Y H,CHAO A L J ,et al.Use of statistical methods to find the polysaccharide structural characteristics and the relationships between monosaccharide composition ratio and macrophage stimulatory activity of regionally different strains of Lentinula edodes[J].Analytica Chimica Acta,2007,584(1):50-56.
[6] KRALOVEC J A,METERA K L,KUMAR J R,et al.Immunostimulatory principles from Chlorella pyrenoidosa-Part 1:Isolation and biological assessment in vitro [J].Phytomedicine,2007,14(1):57-64.
[7] ROEDER A,KIRSCHNING C J,RUPEC R A,et al.Toll-like receptors as key mediators in innate antifungal immunity [J].Medical Mycology,2004,42(6):485-498.
[8] OBERG F,HASEEB A,AHNFELT M,et al.Herbal melanin activates TLR4/NF-kappa B signaling pathway[J].Phytomedicine,2009,16(5):477-484.
[9] PARK H J,HONG J H,KWON H J,et al.TLR4-mediated activation of mouse macrophages by Korean mistletoe lectin-C (KML-C) [J].Biochemical and Biophysical Research Communications,2010,396(3):721-725.
[10] ANDO I,TSUKUMO Y,WAKABAYASHI T,et al.Safflower polysaccharides activate the transcription factor NF-kappa B via Toll-like receptor 4 and induce cytokine production by macrophages[J].International Immunopharmacology,2002.2(8):1155-1162.
[11] HAN SB,YOON Y D,AHN H J,et al.Toll-like receptor-mediated activation of B cells and macrophages by polysaccharide isolated from cell culture of acanthopanax senticosus [J].International Immunopharmacology,2003.3(9):1301-1312.
[12] LI X,W X U.TLR4-mediated activation of macrophages by the polysaccharide fraction from polyporus umbellatus(pers.) fries [J].J Ethnopharmacol,2010,3(6):168-176.
[13] YOON Y D,HAN SB,KANG J S,et al.Toll-like receptor 4-dependent activation of macrophages by polysaccharide isolated from the radix of Platycodon grandiflorum [J].Int Immunopharmacol,2003,3(13-14):1873-1882.
[14] LISA A PRICE,CYNTHIA A WENNER,DANIEL T SLOPER ,et al.Role for toll-like receptor 4 in TNF-alpha secretion by murine macrophages in response to polysaccharide Krestin,a Trametes versicolor mushroom extract[J].Fitoterapia,2010,81(2):914-919.
[15] YIN X L,CHEN L,LIU Y,et al.Enhancement of the innate immune response of bladder epithelial cells by astragalus polysaccharides through upregulation of TLR4 expression[J].Biochemical and Biophysical Research Communications,2010,397(2):232-238.
[16] TAKEUCHI O,S AKIRA.Pattern recognition receptors and inflammation [J].Cell,2010,140(6):805-820.
[17] SCHEPETKIN I A,M T QUINN.Botanical polysaccharides:macrophage immunomodulation and therapeutic potential[J].Int Immunopharmacol,2006,6(3):317-333.
[18] LI B,CRAMER D,WAGNER S,et al.Yeast glucan particles activate murine resident macrophages to secrete proinflammatory cytokines via MyD88-and Syk kinase-dependent pathways[J].Clinical Immunology,2007,124(2):170-181.
[19] ACTON SL,SCHERER P E,LODISH H F,et al.Expression cloning of Sr-Bi,a Cd36-related class-B scavenger receptor[J].Journal of Biological Chemistry,1994,269(33):21003-21009.
[20] IICHMANN A,BURGDORF S,SCHEURER S,et al.Glycation of a food allergen by the maillard reaction enhances its T-cell immunogenicity:Role of macrophage scavenger receptor class a typeⅠand Ⅱ [J].Journal of Allergy and Clinical Immunology,2010,125(1):175-183.
[21] NAKAMURA T,SUZUKI H,WADA,Y,et al.Fucoidan induces nitric oxide production via p38 mitogen-activated protein kinase and NF-kappa B-dependent signaling pathways through macrophage scavenger receptors [J].Biochemical and Biophysical Research Communications,2006,343(1):286-294.
[22] YANGJW,YOON SY,OH SJ,et al.Bifunctional effects of fucoidan on the expression of inducible nitric oxide synthase [J].Biochemical and Biophysical Research Communications,2006,346(1):345-350.
[23] UNDERHILL D M,ROSSNAGLE E,LOWELL C A,et al.Dectin-1 activates Syk tyrosine kinase in a dynamic subset of macrophages for reactive oxygen production [J].Blood,2005,106(7):2543-2550.
[24] BROWN G D.Dectin-1:a signalling non-TLR pattern-recognition receptor[J].Nature Reviews Immunology,2006,6(1):33-43.
[25] HARADA T,N OHNO.Contribution of dectin-1 and granulocyte macrophage-colony stimulating factor(GM-CSF)to mmunomodulating actions of beta-glucan [J].International Immunopharmacology,2008,8(4):556-566.
[26] GUO L,XIE J H,RUAN Y Y,et al.Characterization and immunostimulatory activity of a polysaccharide from the spores of Ganoderma lucidum [J].International Immunopharmacology,2009,9(10):1175-1182.
[27] CHAUNG H C.HUANG T C,YU J H,et al.Immuno-modulatory effects of beta-glucans on porcine alveolar macrophages and bone marrow haematopoietic cell-derived dendritic cells[J].Veterinary Immunology and Immunopathology,2009,131(3-4):147-157.
[28] SAIJO S,FUJIKADO N,FURUTA T,et al.Dectin-1 is required for host defense against Pneumocystis carinii but not against Candida albicans [J].Nature Immunology,2007,8(1):39-46.
[29] GAZI U,L MARTINEZ-POMARES.Influence of the mannose receptor in host immune responses[J].Immunobiology,2009,214(7):554-561.
[30] LIU C,LEUNG M Y K,KOON J C M,et al.Macrophage activation by polysaccharide biological response modifier isolated from Aloe vera L.var.chinensis(Haw.) Berg [J].International Immunopharmacology,2006,6(11):1634-1641.
[31] LI MING,CHUN ,L D S,et al.Effect of ganoderma polysaccharides on cAMP in murine peritoneal macrophages China [J].Journal of Materia Medica,2000,25(1):40-43.
[32] LI MING,CHUN,L L S,et al.Effect of Ganoderma lucidum polysaccharides on inositlo trisphate and diacylglycerol in murineT cells[J].China Pharm J,2001,36(8):522-526.
[33] BELSKA N V,GURIEV A M,DANILETS M G,et al.Water-soluble polysaccharide obtained from acorus calamus L.classically activates macrophages and stimulates Th1 response [J].International Immunopharmacology,2010,10(8):933-942.
[34] WEI S D,YANG K,GONG J P.Effect of GITR/GITRL signaling syetem on macrophage[J].Cell and Molecular immunology,2009,25(12):1207-1209.
[35] CHAUNG H C,HUANG T C,YU J H,et al.Immunomodulatory effects of beta-glucans on porcine alveolar macrophages and bone marrow haematopoietic cell-derived dendritic cells[J].Veterinary Immunology and Immunopathology,2009,131(3-4):147-157.
[36] WANG J,ZUO G,LI J,et al.Induction of tumoricidal activity in mouse peritoneal macrophages by ginseng polysaccharide [J].International Journal of Biological Macromolecules,2010,46(4):389-395.
[37] DERGUNOVA M A, ALEXEENKO T V,ZHANAEVA S Y,et al.Characterization of the novel chemically modified fungal polysaccharides as the macrophage stimulators[J].International Immunopharmacology,2009,9(6):729-733.
Polysaccharides activate signaling pathways of macrophage
MA Zhi-xin,ZHAO Lu-hang
(Department of Biochemistry and Genetics,Zhejiang University School of Mecicine,Hangzhou 310058,China)
Polysaccharides extracted from various sources are natural active substances,which may lead to the activation of macrophage via multiple pathways and mechanisms.This article intends to illustrate the signaling pathways of polysaccharides from plants,fungi,algae and other sources,to indentify the mechanisms on the molecular level,and to explore the novel target immunomodulatory agents.
Polysaccharides;Macrophages/immunol;Signal Transduction
Q 53
A
1008-9292(2011)05-0567-06
http:∥www.journals.zju.edu.cn/med
10.3785/j.issn.1008-9292.2011.05.018
2011-01-18
2011-07-21
馬志新(1982-),男,碩士生,生物化學與分子生物學.
趙魯杭(1962-),男,博士生,副教授,碩士生導師,主要從事糖生物學及免疫學研究;E-mail:zhaoluhang@263.net
[責任編輯 黃曉花]
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
