黃海春季海洋病毒的空間分布特征
2011-09-24 08:11:46殷克東林志芬袁翔城
海洋科學 2011年2期
何 蕾, 殷克東, 林志芬, 田 甜, 袁翔城
(1. 中國科學院 南海海洋研究所熱帶海洋環(huán)境動力學重點實驗室, 廣東 廣州 510301; 2. 中國科學院研究生院, 北京 100049; 3. 中國科學院 海洋研究所海洋環(huán)流與波動重點實驗室, 山東 青島 266071)
黃海春季海洋病毒的空間分布特征
何 蕾1,2,3, 殷克東1, 林志芬1, 田 甜1, 袁翔城1
(1. 中國科學院 南海海洋研究所熱帶海洋環(huán)境動力學重點實驗室, 廣東 廣州 510301; 2. 中國科學院研究生院, 北京 100049; 3. 中國科學院 海洋研究所海洋環(huán)流與波動重點實驗室, 山東 青島 266071)
采用熒光顯微技術(shù), 報道了春季(2006年4月)海洋病毒豐度在黃海的空間分布, 分析了病毒豐度與細菌豐度、葉綠素a濃度、溫度和鹽度的相關(guān)關(guān)系。研究結(jié)果表明, 黃海表層水中病毒豐度的變化范圍為 1.08×107~9.90×107個/mL, 平均值是 5.62×107個/mL±2.28×107個/mL, 細菌豐度的變化范圍為0.01×107~0.16×107個/mL, 平均值是 0.07×107個/mL±0.03×107個/mL, 病毒豐度大于細菌豐度兩個數(shù)量級。葉綠素a濃度表層的變化范圍為0.09~4.94 μg/L, 平均值是0.75 μg/L±0.84 μg/L, 最高值出現(xiàn)在長江口附近。通過相關(guān)性分析, 在黃海春季, 細菌可能控制著病毒的空間分布, 并且病毒的空間分布受鹽度影響較小, 受溫度影響較大。
黃海; 病毒豐度; 細菌豐度; 空間分布
海洋是地球生命的起源地。海洋生命的種類涵蓋了從病毒到高等哺乳類的各個層次。游離的病毒和細菌存在于海水當中已經(jīng)是眾所周知的事情, 自從Azam等[1]科學家們在1983年提出微生物食物環(huán)的概念以來, 浮游細菌的生物學和生態(tài)學研究成為生物海洋學最活躍的前沿領(lǐng)域之一。而在最近20年對海洋病毒的研究中, 人們不但發(fā)現(xiàn)在海洋中病毒數(shù)量是最豐富的[2], 而且還發(fā)現(xiàn)病毒可以感染海洋中大部分的生物[3-5]。這些重大發(fā)現(xiàn)使人們對海洋病毒在生態(tài)環(huán)境方面的重要性有了重新認識, 同時也引起了各國研究學者對海洋病毒的強烈關(guān)注。研究結(jié)果表明海洋病毒是海洋生態(tài)系統(tǒng)中的活躍分子,不僅是微生物食物環(huán)和生物地球化學循環(huán)中的重要角色[2], 還在赤潮控制[6], 基因轉(zhuǎn)移[7]和全球氣候變化[2]等方面起著非常重要的作用。
目前, 國際上已普遍采用熒光顯微技術(shù)對水體中病毒生物量進行檢測[8]。Hara等[9]首次利用熒光顯微計數(shù)法檢測了日本大阪灣和Otsuchi Bay兩個灣域中游離病毒的豐度, 測得病毒豐度為 0.12×107~3.5×107個/mL。Hennes等[10]對墨西哥西部海灣表面水體中的病毒生物量進行檢測, 病毒豐度為 1×107~12×107個/mL。Noble 和 Fuhrman[11]對不同海水中病毒生物量進行計數(shù), 其范圍為 0.03×107~15×107個/mL。He等[12]首次報道了中國南海陸架海區(qū)表層水中病毒豐度為 1.41×107個/mL±0.65×107個/mL。趙苑等[13]對膠州灣浮游病毒進行了研究, 膠州灣浮游病毒的豐度為 0.48×107~22.78×107個/mL 。
黃海是頗為重要的陸架海區(qū), 也是世界上最典型的半封閉性陸架淺海之一。 它與其他陸架海區(qū)一樣, 是人類活動、經(jīng)濟開發(fā)最為集中的地帶, 由于黃海水深較淺, 緊鄰陸地, 多條大河直接或間接注入,所以營養(yǎng)物質(zhì)十分豐富, 生物資源蘊藏量很高, 一向以生產(chǎn)力高而著稱[14]。在這一生態(tài)系統(tǒng)中, 海洋浮游病毒的生態(tài)效應(yīng)如何, 現(xiàn)在并不清楚。本文利用熒光顯微技術(shù)對黃海的病毒豐度進行計數(shù), 并分析病毒豐度與其影響因子的關(guān)系, 探討病毒豐度在黃海生態(tài)系統(tǒng)中的分布特征及其機制, 以期為黃海生態(tài)環(huán)境的研究及改善提供基礎(chǔ)資料。
1 材料與方法
2006年4月“東方紅2號”科學調(diào)查船在黃海進行了海上大規(guī)模調(diào)查, 調(diào)查范圍為北緯 32°31′~39°12′, 東經(jīng) 120°45′~124°, 共設(shè) 41 個站位。(圖 1)
需要測量獲得的參數(shù)有溫度、鹽度、葉綠素、細菌豐度和病毒豐度。使用Sea-Bird SBE CTD 儀器來獲得水體的溫度和鹽度。水樣各采樣點均用 2.5L的Niskin采水器獲得。葉綠素a水樣采樣參照《海洋調(diào)查規(guī)范》(海洋生物調(diào)查和化學調(diào)查), 用Turner Designers 700萃取熒光法測定[15]。
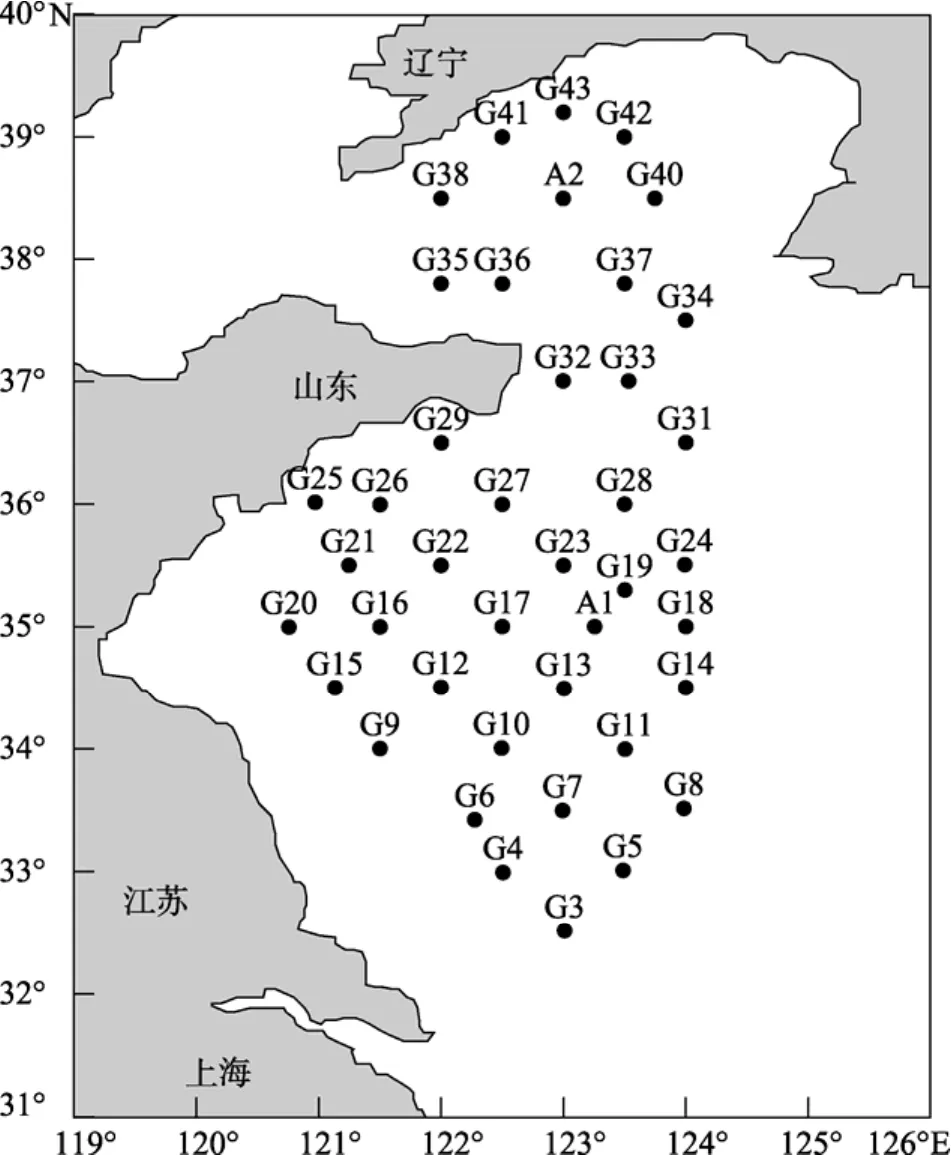
圖1 2006年4月黃海站位Fig. 1 Locations of the sampling stations in the Yellow Sea during April 2006
病毒樣品取 1 mL水樣用戊二醛(終濃度 0.5%)黑暗固定15 min, 然后取0.8 mL樣品過濾到0.02 μm氧化鋁膜(Whatman, Maidstone, England)上, 干燥后,用 0.25% SYBR-Green-I染色, 并在黑暗中放置 15 min, 薄片制好后放在 ?20 oC下保存。樣品在一個月內(nèi)用熒光顯微鏡(OLYMPUS BX41 microscope,×1000 )進行拍照計數(shù)[10]。細菌樣品取10 mL水樣用甲醛(終濃度 5%)固定保存, 回到實驗室用4′6-diamidino-2-phenylindole(DAPI)染色, 用直徑為25 mm, 孔徑0.2 μm的黑色核孔濾膜(black Nucleopore filter)進行抽濾, 將濾膜放在載波片上, 滴一滴浸油(immersion oil, Germany), 蓋上蓋玻片, 再滴少量浸油, 用熒光顯微鏡(OLYMPUS BX41 microscope,×1 000 )進行拍照計數(shù)[16]。每個樣品隨機取至少 10個視野, 計數(shù)每個視野中所有的顆粒。
實驗數(shù)據(jù)用SPSS軟件分析, 所得實驗結(jié)果用平均值±標準誤差表示, 以P< 0.05表示具有顯著性差異。圖形均使用Surfer軟件(Golden software Inc.CO)繪制。
2 結(jié)果

圖2 黃海2006年4月溫度(T)和鹽度(S)表層分布Fig. 2 Horizontal distributions of temperature (T) and salinity (S) in the surface layer of the Yellow Sea during April 2006.Circle symbols indicate the sampling stations
圖2是2006年4月溫度和鹽度表層分布圖, 表層的溫度變化范圍為4.50~13.74℃, 平均值是9.01℃±2.27℃。表層的鹽度變化范圍為 27.87~34.06PSU,平均值是32.20PSU±1.08PSU。從圖2中可以看出溫度隨緯度增加而降低, 南北溫度差異達 9℃, 在蘇北沿岸有一暖水舌向東北延伸。在海域東南部, 有一較強的暖水舌自濟州島西側(cè)海域向北偏西方向伸展,這一暖水舌表征了傳統(tǒng)上認為的黃海暖流。表層鹽度的差異有7PSU。在黃海研究區(qū)的東南部有一較強的高鹽舌自東南向西北伸展。在35°N附近, 主舌鋒偏向西, 分出的另一舌鋒指向北。這一高鹽水舌的分布大致反映了春季黃海暖流的所在位置。在黃海西部, 有3個低鹽中心, 一個位于山東外海, 一個在江蘇外海, 另一個出現(xiàn)在長江口的東北方。
圖3是2006年4月葉綠素a濃度和細菌豐度表層分布圖, 葉綠素a濃度表層的變化范圍為 0.09~4.94 μg/L, 平均值是 0.75 μg/L±0.84 μg/L。葉綠素a濃度在表層有 3個高值區(qū), 其一在黃海研究區(qū)的南部, 近長江口處(4.94 μg/L), 其他兩處在黃海研究區(qū)東部, G18(1.80 μg/L)和 G31(1.98 μg/L)站。在遼寧和山東的南部海域, 葉綠素a濃度最低。細菌豐度的變化范圍為 0.01×107~0.16×107個/mL, 平均值是 0.07× 107個/mL±0.03×107個/mL。細菌豐度在表層分布上差異很大, 有 1個數(shù)量級的差別。北黃海細菌豐度明顯比南黃海細菌豐度高, 最高值在北黃海 G35 站(0.16×107個/mL)和 G40 站(0.13×107個/mL)。
圖4是2006年4月病毒豐度和病毒豐度與細菌豐度的比值(VBR)表層分布圖。病毒豐度的變化范圍為 1.08×107~9.90×107個/mL, 平均值是 5.62×107個/mL±2.28×107個/mL。病毒豐度在平面分布上差異也很大。最大值在南部G10站(9.90× 107個/mL), 其次是G7站(8.85× 107個/mL), 第三大值黃海北部G5站(8.52×107個/mL)。VBR 的變化范圍為 20.93~427.90,平均值是103.52±87.57。VBR的表面分布差異也很大, 最大值東南部 G28(427.90), 最小值在黃海研究區(qū)的東北部G33(20.93)。

圖3 黃海2006年4月葉綠素a(Chla)濃度和細菌豐度(BA)表層分布Fig. 3 Horizontal distributions of chlorophylla(Chla) and bacterial abundance (BA) in the surface layer of the Yellow Sea during April 2006. Circle symbols indicate the sampling stations
3 討論
3.1 病毒豐度計數(shù)方法的選擇
病毒生物量的正確估計對于微生物生態(tài)研究是非常重要的。當前, 海洋病毒豐度的計數(shù)有如下4種方法: 透射電鏡計數(shù)法、熒光顯微鏡計數(shù)法、流式細胞儀計數(shù)法和最大可能數(shù)法。在這4種方法中, 流式細胞儀法、熒光顯微鏡法和最大似然數(shù)法比較簡單易行, 而透射電鏡法則較難操作。透射電鏡是最早也是較為普遍使用的計數(shù)病毒豐度的方法, 第一個高病毒豐度的數(shù)據(jù)就是用透射電鏡法獲得的[17-18]。但透射電鏡法由于采樣的不均勻和低的探測極限等技術(shù)原因而低估病毒豐度[9-11,19-20]。最大似然數(shù)法簡單易行, 但精確度不高。流式細胞儀法是現(xiàn)在比較流行的方法, 可以區(qū)分病毒和細菌, 但對儀器的靈敏度要求高。而熒光顯微鏡法盡管無法消除小的細菌和大的病毒相互重疊所帶來的誤差[21], 但由于該方法的快速, 使之越來越頻繁地用來估計病毒豐度, 而且樣品的制備完全可以在船上完成, 大量樣品使成本更低, 可以收集更多的數(shù)據(jù), 也增加了統(tǒng)計的精確度[22-23]。所以本研究采用熒光顯微鏡法來計數(shù)病毒豐度。

圖4 黃海2006年4月病毒豐度(VA)和病毒細菌比(VBR)表層分布圖Fig. 4 Horizontal distributions of viral abundance (VA) and viruses to bacteria ratio (VBR) in the surface layer of the Yellow Sea during April 2006. Circle symbols indicate the sampling stations
3.2 病毒豐度與其他海域的比較
最近由于報道了在自然水體中游離病毒顆粒的大量存在, 微生物學家用不同的方法計數(shù)病毒在水生環(huán)境中的數(shù)量。Wommanck等[24]總結(jié)了用不同的方法在不同的水生環(huán)境中病毒的數(shù)量, 其變化范圍為104~108個/mL, 在生產(chǎn)力高、富營養(yǎng)的環(huán)境中, 病毒豐度也高; 在空間分布上呈現(xiàn)近岸豐度高、遠岸豐度低的特點。黃海是頗為重要的陸架海區(qū), 一向以生產(chǎn)力高而著稱[14], 病毒豐度為1.08×107~9.90×107個/mL, 平均值是 5.62×107個/mL±2.28×107個/mL。Hara等[9]首次利用熒光顯微計數(shù)法于1987年9月檢測了日本近岸 Osaka灣游離病毒的豐度, 測得病毒豐度為 3.5×107個/mL, Hennes等[10]與 Weinbauer等[19]使用不同的方法分別測墨西哥灣的病毒豐度為1.0×107~12×107個/mL 和 0.03×107~7.9×107個/mL,與我們測得的結(jié)果非常相近。由于黃海是富營養(yǎng)海區(qū), 而南海是貧營養(yǎng)海區(qū), 所以黃海病毒豐度顯著高于南海病毒豐度(0.35×107~2.51×107個/mL)[12]。
3.3 病毒豐度與其影響因素的關(guān)系
病毒是由一個核酸分子(DNA或RNA)與蛋白質(zhì)構(gòu)成的核酸-蛋白質(zhì)的復合體。它雖然具備了生命活動的最基本特征(復制與遺傳), 但不具備細胞的形態(tài)結(jié)構(gòu), 是不“完全”的生命體, 它們的主要生命活動必須在宿主細胞內(nèi)才能表現(xiàn), 因此, 病毒是徹底的寄生物[25]。海洋病毒的宿主細胞主要是浮游植物和海洋細菌。Boehme等與Jiang等報道美國佛羅里達的 Tampa灣病毒豐度與葉綠素a濃度顯著相關(guān),說明浮游植物是病毒的主要宿主, 可能的解釋是富營養(yǎng)水域浮游植物生物量的增加導致了病毒豐度的增加。然而, 在大部分的海洋環(huán)境中, 研究結(jié)果表明病毒豐度與細菌豐度有著顯著的相關(guān)關(guān)系, 說明細菌是病毒的主要宿主, 細菌的豐度控制著病毒豐度[26-37]。從本研究的數(shù)據(jù)分析(表 1)看, 病毒豐度與細菌豐度呈顯著的負相關(guān), 而與葉綠素a濃度沒有顯著的相關(guān)關(guān)系, 說明細菌很有可能是病毒的宿主, 影響著病毒的空間分布。從圖3和圖4中細菌豐度和病毒豐度的表層分布圖看, 細菌豐度少的地方病毒豐度多, 而葉綠素a濃度有高有低。例如在黃海南部, 近長江口海域, 由于受長江徑流影響, 帶來大量的營養(yǎng)鹽, 刺激浮游植物生長, 所以葉綠素a濃度在長江口附近比較高。由于浮游植物生長旺盛, 對細菌的生長有所抑制而對病毒沒有影響, 所以細菌豐度相對較低, 而病毒豐度較高。
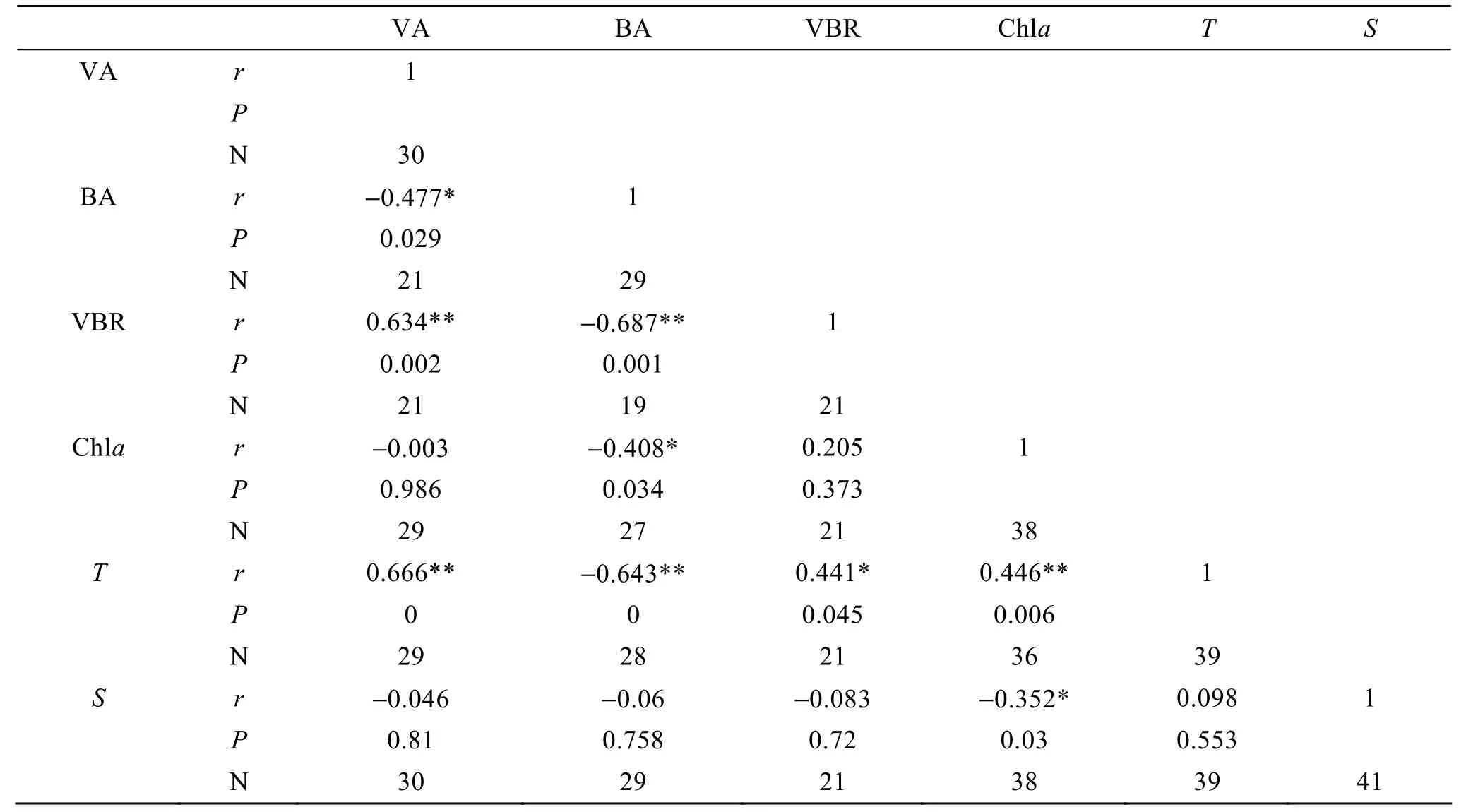
表1 表層病毒豐度與溫度、鹽度、葉綠素a濃度和細菌豐度的相關(guān)關(guān)系Tab. 1 Simple correlation coefficients between virus, bacterial abundance and environmental parameters in the surface layer
海洋細菌與浮游植物的生長繁殖有密切的關(guān)系,一方面, 細菌能夠吸收浮游植物產(chǎn)生的溶解有機物進行二次生產(chǎn)[38], 在為促進自身生長和增殖提供碳源的同時[39], 也為浮游植物的生長提供必要的有機營養(yǎng)和生長因子, 并調(diào)節(jié)浮游植物的微生長環(huán)境[40];另一方面, 細菌可以通過參與生物競爭、分泌特殊物質(zhì)等途徑抑制浮游植物的細胞生長, 甚至裂解其細胞[41]。從表1中可以看出, 葉綠素a濃度與細菌豐度呈顯著的負相關(guān)關(guān)系, 在圖3中, 葉綠素a濃度高的地方細菌豐度低, 說明在黃海浮游植物和細菌的生長有一定的相互抑制作用。
病毒豐度的分布除了受生物因素(浮游植物, 細菌)影響, 同時也受非生物因素(溫度, 鹽度)影響。從表1中可以看出, 病毒豐度、葉綠素a濃度和VBR值與溫度呈顯著的正相關(guān), 細菌豐度與溫度呈顯著的負相關(guān), 說明在春季, 由于太陽輻射, 使表層水增溫, 而且由于緯度的關(guān)系, 越往南增溫越大, 所以合適的溫度條件為生物生長提供條件。由于浮游植物指數(shù)生長時, 抑制細菌生長, 所以病毒就會選擇感染大量的浮游植物, 而使病毒數(shù)量增加, 也就導致了VBR增加。而病毒豐度, 細菌豐度和VBR與鹽度之間沒有顯著的關(guān)系, 說明鹽度對這些參數(shù)的影響并不明顯。根據(jù)以上相關(guān)關(guān)系分析, 黃海海洋病毒的空間分布受鹽度影響較小, 受溫度影響較大。
病毒在海洋生態(tài)系中的重要作用日益受到重視,本文報道了海洋病毒在黃海的空間分布特征, 分析了病毒、細菌、浮游植物、溫度和鹽度的相互影響關(guān)系, 初步探討了海洋病毒在黃海生態(tài)系統(tǒng)中的作用, 為黃海生態(tài)環(huán)境的研究和了解提供資料, 為更全面的研究黃海海洋生態(tài)系統(tǒng)打下基礎(chǔ)。
[1] Azam F, Fenchel T, Field J G, et al. The ecological role of water-column microbes in the sea[J]. Mar Ecol Prog Ser, 1983, 10: 257-263.
[2] Fuhrman J A. Marine viruses and their biologeochemical and ecological and ecological effects[J]. Nature,1999, 399: 541-548.
[3] Bergh O, Borsheim K Y, Bratbak G, et al. High abundance of viruses found in aquatic environments [J].Nature, 1989, 340(6233): 467-468.
[4] Proctor L M, Fuhrman J A. Viral mortality of marine bacteria and cyanobacteria [J]. Nature, 1990, 343:60-62.
[5] Suttle C A, Chan A M, Cottrell M T. Infection of phytoplankton by viruses and reduction of primary productivity[J]. Nature, 1990, 347: 467-469.
[6] Peduzzi P, Weinbauer M G. Effect of concentrating the virus-rich 2 200nm size fraction of seawater on the formation of algal flocs (marine snow) [J]. Limnol Oceanogr, 1993, 38: 1 562-1 565.
[7] Chiura H X. Generalized gene transfer by virus-like particles from marine bacteria[J]. Aquat Microb Ecol,1997, 13: 75-83.
[8] 李洪波, 肖天, 林鳳翱. 海洋浮游病毒的研究方法[J].海洋科學, 2010, 34(9): 97-101.
[9] Hara S, Terauchi K, Koike I. Abundance of viruses in marine waters assessment by epifluorescence and transmission electron microscopy[J]. Appl Environ Microb, 1991, 57: 2 731-2 734.
[10] Hennes K P, Suttle C A, Chan A M. Fluorescently labeled virus probes show that natural virus population can control the structure of marine microbial communities[J]. Appl Environ Microb, 1995, 61: 3 623-3 627.
[11] Noble R T, Fuhrman J A. Use of SYBR Green I for rapid epifluorescence counts of marine viruses and bacteria[J]. Aquat Microb Ecol, 1998, 14: 113-118.
[12] He L, Yin K, Yuan X, et al. Spatial Distribution of Viruses, Bacteria and Chlorophyll in the Northern South China Sea[J]. Aquat Microb Ecol, 2009, 54(2):153-162.
[13] 趙苑, 武洪慶, 李洪波,等. 膠州灣浮游病毒的分布研究[J]. 海洋科學, 2010, 34(7): 86-92.
[14] 王保棟, 劉峰, 戰(zhàn)閏.黃海生源要素的生物地球化學研究評述[J]. 黃渤海海洋, 2001, 19(2): 99-106.
[15] Parsons T R, Maita Y, Lalli C M. A manual of chemical and biological methods for seawater analysis[M]. Oxford: Pergamon Press, 1984: 7-37.
[16] Kepner R L JR, Pratt J R. Use of fluorochromes for direct enumeration of total bacteria in environmental samples: past and present[J]. Microbiol Rev, 1994, 58:603-615
[17] Torrella F, Morita R Y. Evidence for a high incidence of bacteriophage particles in the waters of Yaquina Bay,Oregon: Ecological and taxomomical implications[J].Appl Environ Microb, 1979, 37: 774-778.
[18] Ewert D, Paynter M. Enumeration of bacteriophages and host bacteria in sewage and the activated-sludge treatment process[J]. Appl Environ Microb, 1980, 39:67-69.
[19] Weinbauer M G, Suttle C A. Comparison of epifluorescence and transmission electron microscopy for counting viruses in natural marine waters[J]. Aquat Microb Ecol, 1997, 13: 225-232.
[20] Bettarel Y, Sime-Ngando T, Amblard C, et al. A comparison of methods for counting viruses in aquatic systems[J]. Appl Environ Microb, 2000, 66: 2 283-2 289.
[21] Sommaruga R, Krossbacher M, Salvenmoser W, et al.Presence of large virus-like particles in a eutrophic reservoir[J]. Aquat Microb Ecol, 1995, 9: 305-308.
[22] Proctor L. Advances in the study of marine viruses[J].Microsc Res Tech, 1997, 37: 136-161.
[23] Fuhrman J. Impact of viruses on bacterial processes[C]//Kirchman D. Microbial Ecology of the Oceans. New York: Wiley, 2000: 327-350.
[24] Wommack K E, Colwell R R. Virioplankton: viruses in aquatic ecosystems[J]. Microbiol Mol Biol Rev, 2000,64: 69-114.
[25] 翟中和, 王喜忠, 丁明孝. 細胞生物學[M].北京:高等教育出版社,2000.
[26] Boehme J, Frischer M E, Jiang S C, et al. Viruses, bacterioplankton in the southeastern Gulf of Mexico: distribution and contribution to oceanic DNA pools[J].Mar Ecol Prog Ser, 1993, 97: 1-10.
[27] Cochlan W P, Wikner J, Steward G F, et al. Spatial distribution of viruses, bacteria, and chlorophyll a in neritic, oceanic and estuarine environments[J]. Mar Ecol Prog Ser, 1993, 92: 77-87.
[28] Hara S, Koike I, Terauchi K, et al. Abundance of viruses in deep oceanic waters[J]. Mar Ecol Prog Ser,1996, 145: 269-277.
[29] Hennes K P, Simon M. Significance of bacteriophages for controlling bacterioplankton growth in a mesotrophic lake[J]. Appl Environ Microb, 1995, 61: 333-340.
[30] Jiang S C, Paul J H. Seasonal and diel abundance of viruses and the occurrence of lysogeny/ bacteriocinogeny in the marine environment[J]. Mar Ecol Prog Ser, 1994, 104: 163-172.
[31] Kepner R L, Wharton R A, Suttle C A. Viruses inAntarctic lakes[J]. Limnol Oceanogr, 1998, 43:1 754-1 761.
[32] Paul J H, Rose J B, Jiang S C, et al. Distribution of viral abundance in the reef environment of Key Largo,Florida[J]. Appl Environ Microb, 1993, 59: 718-724.
[33] Steward G F, Smith D C, Azam F. Abundance and production of bacteria and viruses in the Bering and Chukchi Sea[J]. Mar Ecol Prog Ser, 1996, 131: 287-300.
[34] Tuomi P, Torsvik T, Heldal M, et al. Bacterial population dynamics in a meromictic lake[J]. Appl Environ Microb, 1997, 63: 2 181-2 188.
[35] Weinbauer M G, Fuks D, Peduzzi P. Distribution of viruses and dissolved DNA along a coastal trophic gradient in the northern Adriatic Sea[J]. Appl Environ Microb, 1993, 59: 4 074-4 082.
[36] Weinbauer M G, Peduzzi P. Significance of viruses versus heterotrophic nanoflagellates for controlling bacterial abundance in the northern Adriatic Sea[J]. J Plankton Res, 1995, 17: 1 851-1 856.
[37] Weinbauer M G, Fuks D, Puskaric S, et al. Diel, seasonal, and depth-related variability of viruses and dissolved DNA in the Northern Adriatic Sea[J]. Microb Ecol, 1995, 30: 25-41.
[38] 鄭天凌, 王海黎, 洪華生. 微生物在碳的海洋生物地球化學循環(huán)中的作用[J]. 生態(tài)學雜志, 1994, 13(4):47-50.
[39] Jennifer C, James A A E, Ellen T M. Utilization and turnover of labile dissolved organic matter by bacterial heterotrophs in eastern North Pacific surface waters [J].Mar Ecol Prog Ser, 1996, 139: 267-279.
[40] Cho B C, Azam F. Major role of bacteria in biogeochemical fluxes in the ocean's interior [J]. Nature, 1988,332: 441-443.
[41] 周玉航, 潘建明, 葉瑛, 等. 細菌、病毒與浮游植物相互關(guān)系及其對海洋生物地球化學循環(huán)的作用[J].臺灣海峽, 2001, 20(3): 340-345.
Received: Nov., 20, 2009
Key words:Yellow Sea; Viral abundance (VA); Bacterial abundance (BA); spatial distribution
Abstract:The spatial distribution of viral and bacterial abundance as well as chlorophylla(chla) concentrations were investigated during spring (April 2006) in the Yellow Sea. The viral abundance varied from 1.08×107/mL to 9.90×107/mL with the average of 5.62×107/mL±2.28×107/mL in spring, exceeding bacterial abundance (from 0.01×107/mL to 0.16×107/mL with the average of 0.07×107/mL±0.03×107/mL) by two order of magnitude. The chlorophyllaconcentrations varied from 0.09μg/L to 4.94 μg/L with the average of 0.75μg/L±0.84μg/L, and the highest concentration occurred near the Changjiang River estuary. A multiple regression analysis showed that the bacteria and temperature were the main factors governing the viral spatial distribution in the Yellow Sea.
(本文編輯:張培新)
Spatial distribution of viruses in spring in the Yellow Sea
HE Lei1,2,3, YIN Ke-dong1, LIN Zhi-fen1, TIAN Tian1, YUAN Xiang-cheng1
(1. Key Laboratory of Tropical Marine Environmental Dynamics, South China Sea Institute of Oceanology,Chinese Academy of Sciences, Guangzhou 510301, China; 2. Graduate School of the Chinese Academy of Sciences, Beijing 100049, China; 3. Key Laboratory of Ocean Circulation and Waves, Institute of Oceanology,Chinese Academy of Sciences, Qingdao 266071, China)
Q938
A
1000-3096(2011)02-0010-07
2009-11-20;
2010-04-11
國家自然科學基金重大項目(40490264); 國家自然科學基金面上項目(40676020)
何蕾(1976-), 女, 黑龍江省大慶市人, 博士生, 主要從事海洋生態(tài)學研究, E-mail: leihe@scsio.ac.cn
