湖北省棉花主產區煙粉虱生物型分布及系統發育分析
2011-08-27 00:40:46周新改馬偉華劉美剛楊小紅
植物保護 2011年5期
關鍵詞:生物
周新改, 馬偉華*, 劉美剛, 楊小紅, 王 沫
(1.湖北省昆蟲資源利用與害蟲可持續治理重點實驗室,華中農業大學,武漢 430070;2.湖北省仙桃市植物保護站,仙桃 433000; 3.湖北省潛江市植物保護站,潛江 433104)
湖北省棉花主產區煙粉虱生物型分布及系統發育分析
周新改1, 馬偉華1*, 劉美剛2, 楊小紅3, 王 沫1
(1.湖北省昆蟲資源利用與害蟲可持續治理重點實驗室,華中農業大學,武漢 430070;2.湖北省仙桃市植物保護站,仙桃 433000; 3.湖北省潛江市植物保護站,潛江 433104)
[目的]明確武漢、荊州、孝感、隨州、武穴5個棉花主產區煙粉虱生物型組成。[方法]通過以線粒體細胞色素氧化酶I(mitochondrial cytochrome oxidase I,mtCOI)為基礎的PCR-RFLP和基因測序的方法,鑒定湖北5個棉花主產區煙粉虱地理種群的生物型組成,并分析5個地理種群與其他省種群的系統發育關系。[結果]Q型煙粉虱為湖北省棉花主產區的優勢種群,而武漢溫室種群則均為B型煙粉虱,土著生物型ZHJ1型在大田中略有分布;系統發育分析表明,湖北省棉花主產區B型和Q型煙粉虱種群均與中國其他多個省的煙粉虱種群親緣關系較近。[結論]本文研究結果可以為湖北省5個棉花主產區煙粉虱的有效防治提供參考。
煙粉虱; 生物型; PCR-RFLP; 系統發育分析
* 通信作者 E-mail:weihuama@mail.hzau.edu.cn 2006年在武漢地區大發生,煙粉虱由原來的次要害蟲迅速上升為主要害蟲[3]。目前,雖然已有報道稱Q型煙粉虱在我國長江流域已成為優勢種群[1],但湖北省主要棉區煙粉虱生物型組成仍不明了。此外,外來入侵生物的系統發育關系研究對于闡明生物入侵途徑、尋找有效天敵、了解生物入侵后生物學特性變化以及提出有效控制策略均有重要意義[4]。本文通過VspI-mtCOIPCR-RFLP分子標記技術,對湖北省主要5個棉花產區的煙粉虱地理種群生物型進行鑒定,以明確武漢、荊州、孝感、隨州、武穴5個棉花主產區煙粉虱生物型組成,并構建系統發育樹,分析湖北棉花主產區煙粉虱種群與其他省種群的親緣關系。
1 材料與方法
1.1 供試蟲源
供試種群分別采自湖北五大棉花主產區,其寄主均為棉花‘鄂棉23’,地理種群、采集地、生物型、采集時間、樣本量及GenBank查詢碼見表1。

表1 湖北省棉花上煙粉虱種群及生物型鑒定
1.2 單頭煙粉虱DNA提取
參照 Kirk等[5]和 Frohich等[6]方法,略有改動。取單頭煙粉虱在ddH2O中清洗,置于含有60μL堿裂解液(50mmol/L,Tris-HCl(pH8.0),20mmol/L NaCl,1.0mmol/L EDTA,1%SDS)的1.5mL離心管中,用勻漿器充分勻漿后,在55℃水浴鍋中溫育1h。取出后,加入2倍體積的預冷無水乙醇,冰浴2h。12 000r/min離心10min,去除上清液,70%乙醇清洗沉淀,再次12 000r/min離心10min,棄上清液后室溫風干,ddH2O溶解后即可用于PCR反應。
1.3 PCR擴增及電泳
所用引物 C1-J-2195(5′-TTGATTTTTTGGTCATCCAGAAGT-3′),tRNA-1576 (5′-TATAAATCTTAAAACTGCA-3′)[7]。擴增出大小約為800 bp的PCR產物。PCR反應體系總體積為50μL,含2.5UTaq酶,引物C1-J-2195和tRNA-1576(10μmol/L)各2μL,模板DNA 2μL。反應體系于94℃預變性5min后,進行35個循環:94℃ 變性1min;50℃退火1min;72℃延伸1min。循環結束后72℃ 延伸5min。PCR產物于1%的瓊脂糖凝膠電泳分離,EB染色后于凝膠成像系統下觀察。
1.4 VspI-mtCOI PCR-RFLP鑒定生物型
PCR產物直接用于限制性內切酶VspI(特異性識別位點:ATTAAT)酶切反應,酶切的方法和步驟完全參照酶產品說明書(Ferment Inc.,Glen Burnie,MD)。酶切后于1%的瓊脂糖凝膠電泳分離,EB染色后于凝膠成像系統下觀察。B型煙粉虱個體的mtCOI基因片段酶切后為大小約700bp的單一條帶,而Q型煙粉虱個體的mtCOI基因片段酶切后為大小約500bp和300bp的2條譜帶[8-10]。
1.5 序列測定與數據分析
PCR產物純化后送華大基因公司測序,測序結果提交至 GenBank(http:∥www.ncbi.nlm.nih.gov)。得到的測序結果用Clustal W進行排序分析,比較所得序列之間的差異,并與GenBank中煙粉虱mtCOI部分序列進行比較。以溫室白粉虱(Trialeurodes vaporariorum)相應的mtCOI基因片段序列為外群,通過p-distance模型構建系統發育聚類樹(MEGA 3.0),系統發育樹各分支的置信度(boostrap)均進行2 000次重復檢驗。
2 結果與分析
2.1 PCR-RFLP鑒別湖北五大棉花主產區煙粉虱地理種群生物型
PCR擴增產物的瓊脂糖凝膠電泳結果表明,每個樣品的擴增產物約850bp(圖1)。由圖1可見,武漢、孝感、荊州、武穴和隨州大田的煙粉虱種群的VspI酶切產物都會在電泳后顯示兩條可見譜帶,一條大小約為500bp,另一條大小約300bp,而武漢溫室的兩個煙粉虱種群VspI酶切產物都會在電泳后顯示1條大小約700bp的可見譜帶。根據VspI-mtCOI PCR-RFLP鑒定生物型結果,采自武漢、孝感、武穴、隨州和荊州大田的煙粉虱種群都為Q型,而采自武漢溫室的煙粉虱種群均為B型(圖1)。總體來看,湖北棉花主產區大田煙粉虱種群以Q型為主,而溫室種群則均為B型煙粉虱。

圖1 煙粉虱mtCOI DNA PCR產物及VspI酶切產物膠片圖
2.2 系統發育分析
由圖2可見,采自溫室的煙粉虱種群與福建、云南、廣東、廣州、海南、山西、北京、內蒙古、河南、浙江以及山東的B型煙粉虱聚為同一進化支,而ZHJ型和外族溫室白粉虱各聚為一進化支,說明湖北武漢溫室B型煙粉虱與多個省份的B型種群親緣關系較近。

圖2 中國B型、ZHJ型煙粉虱系統發育樹
與B型煙粉虱相似,采自湖北武漢、隨州、荊州、孝感的煙粉虱種群以及武穴種群的大部分個體與多個省份的Q型種群親緣關系較近,且所有Q型個體都處于同一分支(圖3)。但是武穴種群的一部分煙粉虱個體(WuXue09-2,HM597858;WuXue09-4,HM597860;WuXue09-5,HM597861;2009年10月鑒定,表1,圖3)與中國浙江的ZHJ1型種群聚為同一進化支,這不僅說明武穴的煙粉虱種群由Q型和ZHJ1型兩個生物型組成(圖3),而且也說明VspI-mtCOIPCR-RFLP技術在區分Q型和ZHJ1型煙粉虱時存在缺陷。2個月后,再次檢測武穴品系生物型組成,則發現所有被測個體都為Q型煙粉虱(WuX-ue09-6,HM626150;WuXue09-7,HM626151;WuX-ue09-8,HM626152;WuXue09-7,HM626151;2009年12月鑒定,表1,圖3),說明在室內飼養條件下,Q型煙粉虱的種群競爭能力優于土著ZHJ1型煙粉虱。
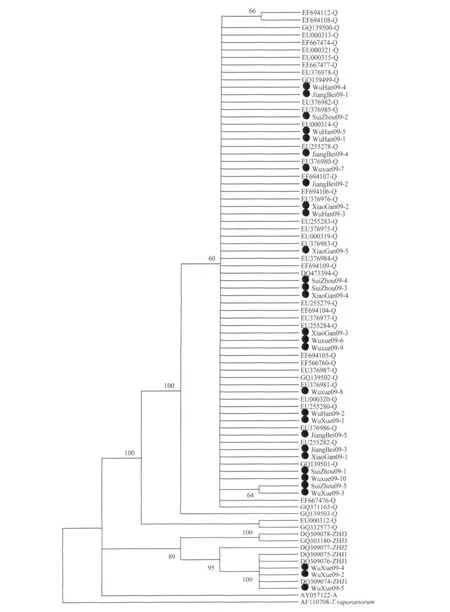
圖3 中國Q型、ZHJ型煙粉虱系統發育樹
3 討論
煙粉虱生物型種類眾多,除B型和Q型煙粉虱外,大部分生物型都分布在特定的區域。B型和Q型煙粉虱均是近20年來世界上分布范圍最廣的外來入侵物種之一,并分別在20世紀末和21世紀初侵入我國,使我國棉花、蔬菜及觀賞植物等種植業蒙受巨大經濟損失[11-12]。目前,煙粉虱的防治措施主要還是依靠化學防治,據報道,B型和Q型煙粉虱對藥劑的敏感性往往不同,且不同生物型煙粉虱對寄主植物的適應性不同[13],因此,明確煙粉虱地理種群的生物型組成,對制定合理有效的防治措施意義重大。本文通過VspI-mtCOIPCR-RFLP和mtCOI序列分析的方法,在湖北省五大棉花主產區共鑒定出3種煙粉虱生物型,它們分別是B型(武漢溫室)、Q型(武漢、隨州、武穴、荊州及孝感大田)和ZHJ1型(武穴大田)(表1)。其中,Q型煙粉虱分布最廣,在所有采集地均有分布,為大田優勢種群;B型煙粉虱僅在溫室中有分布,且在溫室中沒有發現Q型個體;ZHJ1型僅在武穴棉區有少量分布,是唯一一個土著煙粉虱種群。萬方浩等曾報道,Q型煙粉虱為長江流域的優勢種群[1],與本文結果一致。此外,分別于2008、2009年2次自溫室采集到的煙粉虱種群不僅均為B型煙粉虱,且同源性極高(圖2),說明B型煙粉虱種群在溫室中能穩定存在,并可能常年發生,而不受Q型煙粉虱影響,與大田情況相反。總之,湖北省Q型煙粉虱在棉花大田條件下占絕對優勢,而B型煙粉虱則在溫室條件下優勢較明顯,土著生物型ZHJ1型在大田中也有零星分布。
系統發育分析表明,湖北省武漢溫室B型和棉花主產區Q型煙粉虱均與多個周邊省份煙粉虱種群親緣關系較近(圖2,3),說明入侵湖北的煙粉虱種群主要來源于國內種群遷入,而并非國外入侵種群直接侵入。此外,無論B型和Q型煙粉虱,所有被檢測個體在系統發育樹上都處于同一進化支,由此可以推測,中國多省B型或Q型煙粉虱不同地理種群間的遺傳差異較小,這說明中國多省煙粉虱地理種群間基因交流較頻繁,無明顯地域阻隔。
武穴煙粉虱種群一部分與浙江的ZHJ1型聚為同一進化支,這不僅說明武穴的煙粉虱種群由Q型和ZHJ1型2個生物型組成(表1,圖3),而且也說明VspI-mtCOIPCR-RFLP技術在區分 Q 型 和ZHJ1型煙粉虱時存在缺陷。據報道,PCR-RFLP技術是當前眾多煙粉虱生物型分子鑒定技術中操作最簡單、經濟代價最低、應用最廣泛的技術之一,且已有報道證明VspI-mtCOIPCR-RFLP技術可適用于鑒別世界各地的B型和Q型煙粉虱種群,而本文的發現卻說明,該技術在用于鑒別Q型和土著生物型ZHJ1型煙粉虱時存在缺陷,需要后續研究尋找出更精確的煙粉虱生物型鑒定技術。
武穴煙粉虱種群在室內飼養2個月后的生物型組成為40%Q型和60%ZHJ1型(表1,圖3),說明武穴種群在田間為Q型和ZHJ1型2個生物型組成的混合種群,且兩生物型間的比例接近1∶1。但在室內飼養4個月后,該種群則變為Q型煙粉虱種群(表1,圖3),說明ZHJ1型和Q型煙粉虱存在種群競爭現象,且在室內飼養條件下,Q型煙粉虱的種群競爭能力明顯優于ZHJ1型煙粉虱,而與大田條件下的情況不同。但兩生物型間的種群競爭機理尚不清楚,需要開展進一步的研究。
[1] 萬方浩,張桂芬,呂志創,等.B型煙粉虱的入侵機理與控制基礎[J].中國科學,2009,3(2):141-148.
[2] 饒瓊,張宏宇,羅晨,等.武漢地區煙粉虱的危害及其生物型鑒定[J].華中農業大學學報,2009,28(5):535-539.
[3] 周國珍,董長珊.湖北省蔬菜煙粉虱發生與危害初步調查[J].湖北植保,2006(2):19-20.
[4] Tsutsui N D,Suarez A V,Holway D A,et al.Relationships among native and introduced populations of the Argentine ant(Linepithema humile)and the source of introduced populations[J].Molecular Ecology,2001,10(9):2151-2161.
[5] Kirk A A,Lacey L A,Brown J K,et al.Variation in theBemisia tabacis.l.species complex (Hemiptera:Aleyrodidae)and its natural enemies leading to successful biological control ofBemisia tabacibiotype B in the USA[J].Bulletin of Entomological Research,2000,90:317-327.
[6] Frohlich D R,Torres J I,Bedford I D,et al.A phylogeographical analysis of theBemisia tabacispecies complex based on mitochondrial DNA markers[J].Molecular Ecology,1999,8(10):1683-1691.
[7] Zang L S,Chen W Q,Liu S S.Comparison of performance on different host plants between the B biotype and a non-B biotype ofBemisia tabacifrom Zhejiang,China[J].Entomologia Experimentalis et Applicata,2006,121:221-227.
[8] 林克劍,吳孔明,魏洪義.煙粉虱在不同寄主作物上的種群動態及化學防治[J].昆蟲知識,2002,39(4):284-288.
[9] Horowitz A R,Kontsedalov S,Khasdan V,et al.Biotypes B and Q ofBemisia tabaciand their relevance to neonicotinoid and pyriproxyfen resistance[J].Archives of Insect Biochemistry and Physiology,2005,58:216-225.
[10]Khasdan V,Levin I,Rosner A,et al.DNA markers for identifying biotypes B and Q ofBemisia tabaci(Homoptera:Aleyrodidae)and studying population dynamics[J].Bulletin of Entomological Research,2005,95:605-613.
[11]褚棟,張友軍,叢斌,等.世界性重要害蟲B型煙粉虱的入侵機制[J].昆蟲學報,2004,47(3):400-406.
[12]劉樹生,張友軍,羅晨,等.煙粉虱[M]∥萬方浩,鄭小波,郭建英.重要農林外來入侵物種的生物學與控制.北京:科學出版社,2005:69-128.
[13]Iida H,Kitamura T,Honda K.Comparison of egg-hatching rate,survival rate and development time of the immature stage between B and Q biotypes ofBemisia tabaci(Gennadius)(Homoptera:Aleyrodidae)on various agricultural crops[J].Applied Entomology and Zoology,2009,44(2):267-273.
Biotype distribution and phylogenetic analysis ofBemisia tabacipopulations in main cotton planting areas in Hubei Province
Zhou Xingai1, Ma Weihua1, Liu Meigang2, Yang Xiaohong3, Wang Mo1
(1.Hubei Key Laboratory of Insect Resources Utilization and Sustainable Pest Management,Huazhong Agricultural University,Wuhan430070,China; 2.Xiantao Station of Plant Protection,Hubei433000,China;3.Qianjiang Station of Plant Protection,Hubei433104,China)
[Objective]To understand the biotypes ofBemisia tabaciof cotton in main planting areas of Wuhan,Jingzhou,Xiaogan,Suizhou and Wuxue in Hubei Province.[Method]In this study,PCR-RFLP of mtCOI and gene sequencing method were used to identify the biotypes of whitefly field populations from main cotton planting areas in Hubei Province,and phylogenetic analysis of them was also conducted.[Result]The Q biotype was dominant in the field populations,while the B biotype was dominant in greenhouse populations.In addition,ZHJ1 biotype was also identified in Wuxue population.Phylogenetic analysis showed that populations from main cotton planting areas in Hubei Province were always homologous with populations from most other provinces in China.[Conclusion]The results may provide a reference for the control ofB.tabaciin Hubei Province.
Bemisia tabaci; biotype; PCR-RFLP; phylogenetic analysis
公益性行業(農業)科研專項(200803005);植物病蟲害生物學國家重點實驗室開放基金(SKL2010OP03);華中農業大學新教師經費(5220408080,5220409004)
S 435.622.9
A
10.3969/j.issn.0529-1542.2011.05.012
煙粉虱[Bemisia tabaci(Gennadius)]又名棉粉虱、甘薯粉虱、銀葉粉虱,在世界各地均有分布。其危害包括直接吸取植物汁液,分泌蜜露影響植物的光合作用,以及傳播植物病毒,給農業生產造成了巨大的經濟損失。煙粉虱是一個正處于快速進化過程的復合種(species complex),截至2005年,煙粉虱已有26個種群被賦予特定生物型,包括A、AN、B、B2、C、Cassava(木薯)、D、E、F、G、G/H、H、I、J、K、L、M、N、NA、Okra (秋葵)、P、Q、R、S、ZHJ、FJ型[1],不同的煙粉虱生物型在寄主范圍、傳毒能力、地理分布及抗藥性等方面存在顯著差異[2]。
在湖北省,煙粉虱一直作為次要害蟲存在,但于
2010-11-03
2010-11-25
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
