Ipr1對巨噬細胞抗結核分枝桿菌感染免疫相關基因表達的影響*
2011-08-21 10:23:28劉鵬飛李波清喬媛媛張玉梅
中國人獸共患病學報 2011年8期
關鍵詞:檢測
李 娜,劉鵬飛,李波清,喬媛媛,張玉梅
2.濱州醫學院附屬醫院神經外科,煙臺 264003
Ipr1對巨噬細胞抗結核分枝桿菌感染免疫相關基因表達的影響*
李 娜1,劉鵬飛2,李波清1,喬媛媛1,張玉梅1
目的應用基因芯片技術檢測Ipr1的表達對巨噬細胞感染分枝桿菌H37Ra后參與免疫應答的相關基因的表達差異。方法實驗組與對照組細胞分別感染分枝桿菌H37Ra,基因芯片檢測實驗組和對照組細胞感染H37Ra 96h后免疫相關基因表達差異。定量PCR反應檢測芯片結果中上調的3個基因的表達差異用以驗證芯片結果的可靠性。結果基因芯片檢測結果顯示Ipr1基因的表達上調了11個固有免疫機制中相關基因表達,其中與MΦ抗Mtb感染關系密切的基因有:TLR2、TLR4、Irak1、Traf6、Ifngr1、Tnfrsf1a,而參與調節適應性免疫應答的相關基因如:IL-1,IL-12無明顯表達差異。結論
Ipr1增強MΦ抗Mtb感染的機制可能主要是通過上調MΦ抗感染固有免疫機制的有關基因的表達從而發揮抗菌作用。本實驗為進一步研究Ipr1促進MΦ的活化及殺傷胞內吞噬的Mtb的作用機制奠定了基礎。
Ipr1;結核分枝桿菌;巨噬細胞
結核病是嚴重威脅人類健康的疾病,全球每年約有800萬新發病例,170多萬人死亡,死亡數居單一傳染病之首[1]。結核分枝桿菌(Mycobacterium tuberculosis,Mtb)為細胞內寄生菌。越來越多的研究表明固有免疫應答在抗結核分枝桿菌感染中發揮著重要的作用。而巨噬細胞(MΦ)是固有免疫應答中最主要的細胞,它是機體抗Mtb感染的第一道防線[2-3],2005年,Pan等在小鼠MΦ內發現了一個介導MΦ抗胞內病原體的基因—胞內病原體抗性基因1(intracellular pathogen resistance 1,Ipr1),該基因具有體外調節MΦ對Mtb反應能力的作用[4]。
本實驗室已成功構建Ipr1基因真核表達基因pEGFP-Ipr1并且驗證Ipr1基因產物表達部位位于細胞核內[5]。在本研究中將利用基因芯片技術研究Ipr1基因的表達與M Φ抗分枝桿菌感染固有免疫相關機制之間是否有著某種相互關聯,從而為闡明Ipr1在MΦ抗結核病固有免疫中的機制奠定實驗基礎。
1 材料與方法
1.1 質粒、細胞和菌株 質粒pEGFP-Ipr1由本實驗室構建保存,質粒pEGFP-C1、細胞株RAW264.3由本實驗室保存。分枝桿菌減毒株H37Ra為北京藥品生物制品檢定所提供(菌號93020)。
1.2 主要試劑 RPMI1640培養基干粉、1∶125胰蛋白酶(美國Gibco公司);6孔細胞培養板、24孔細胞培養板(美國Costar公司);胎牛血清(杭州四季青公司);真核細胞轉染試劑 Lipofectamine2000(Invitrogen公司);M7H9培養基干粉、M7H10培養基干粉及OADC enrichment(Difco公司)。美國SuperArray公司的小鼠固有免疫和適應性免疫應答基因芯片(Oligo Innate&Adaptive Immune Responses Microarray)購自上海康成公司。
1.3 穩定表達 Ipr1的 RAW264.7篩選RAW264.7細胞分別轉染pEGFP-Ipr1及空質粒pEGFP-C1,G418篩選穩定表達Ipr1的RAW264.7細胞及穩定表達綠色熒光蛋白的 RAW264.7細胞[6],分別命名為實驗組及對照組。
1.4 結核分枝桿菌H37Ra的培養及細菌懸液的準備 挑取改良羅氏培養基上處于對數生長期的新鮮分枝桿菌H37Ra菌落,以0.05%吐溫鹽水充分研磨后,接種M7H9液體培養基,37℃靜止培養約3 w后,密閉離心棄上清,以高壓滅菌的0.9%生理鹽水反復淘洗后自然沉淀3次,每次10min,取上清,無血清RPMI1640培養液重新調整細菌濃度為1×107/mL制成細菌懸液備用。
1.5 分枝桿菌H37Ra感染巨噬細胞及細胞吞噬細菌量檢查 經G418篩選得到的實驗組細胞與對照組細胞常規用含10%胎牛血清、600μ g/mL G418的RPMI 1 640培養液在 5%CO2、37℃、飽和濕度條件下培養。待細胞狀態良好,將實驗組細胞和對照組細胞分別用0.25%的胰蛋白酶消化并收獲細胞,轉種至6孔板,每孔細胞數為1×106,同時常規制備6孔板細胞爬片用于抗酸染色。繼續用含10%胎牛血清、600μ g/mL G418的 RPMI1640培養基培養,待細胞貼壁換用無G418、無抗生素的含10%胎牛血清 RPMI1640培養基培養細胞,同時加入PMA,使其終濃度為 100nmol/L。24h后,將前述準備的分枝桿菌H37Ra細菌懸液按細胞∶細菌=1∶10的比例加入到6孔板的實驗組和對照組細胞培養孔內,5%CO2、37℃培養6h,用含1%胎牛血清的PBS洗滌3次,洗去未被MΦ吞噬的細菌,換入新鮮的不含抗生素的完全RPMI1640繼續培養(為便于描述將此時間點計為0h)。之后每隔6h將細胞用含1%胎牛血清的PBS洗滌3次,換入新鮮不含抗生素的完全RPMI1640繼續培養。24h后將細胞常規洗滌3次取出實驗組和對照組細胞爬片進行抗酸染色觀察MΦ吞噬細菌情況。
1.6 基因芯片檢測 細菌感染96h后,抽提6孔板中各組細胞總RNA。提取的的總RNA標本經瓊脂糖凝膠電泳檢測完整性及紫外分光光度計檢測OD值以確定其純度及濃度。按相關試劑盒說明進行cDNA的合成及線性生物素標記的cRNA的合成、擴增及純化。按Oligo Innate&Adaptive Immune Responses Microarray基因芯片檢測說明書進行芯片雜交。應用化學發光檢測試劑盒檢測芯片雜交結果。X射線膠片曝光在膠片上顯示檢測結果。X-射線膠片曝光后,將膠片上的圖象用Microtek 120TF掃描儀(上海中晶)掃描并轉換為灰度TIFF格式的圖片文件保存。使用配套軟件GEArray Expression Analysis Suite對原始數據進行完整的芯片數據分析。計算待檢基因表達水平占該看家基因表達水平的百分率然后比較對照組及實驗組特定基因表達水平的差異。
1.7 Real-time PCR檢測 為驗證芯片檢測結果的可靠性選取上述基因芯片檢測結果中上調的3個基因進行Real-time PCR檢測。應用引物設計軟件Primer 5,分別設計了3個待檢基因及內參GAPDH的上下游引物:GAPDH F 5′AAGAAGGTGGTGAAGCAGGC 3′R 5′TCCACCACCCTGT TGCTGTA 3′;Ifngr1 F 5′CT TTGACGAGCACTGAGGA 3′R 5′CCAGGAACCCGAATACACC 3′;Tlr2 F 5′TGTCGT TCAAGGAGGTGCG 3′R 5′TCCAGAAGAGCCAAAGAGC 3′;Tlr4 F 5′GAGAATCTGGTGGCTGTGG 3′R 5′T TCCCTGAAAGGCT TGGTC 3′。采用上述方法進行細菌培養及感染細胞,RNA抽提及 RT-PCR合成cDNA方法同基因芯片檢測。制備用于繪制標準曲線的梯度稀釋DNA模板:針對每一需要測量的基因和管家基因,選擇一確定表達該基因的cDNA模板進行PCR反應,PCR產物與100 bp DNA Ladder在2%瓊脂糖凝膠電泳,溴化乙錠染色,檢測PCR產物是否為單一特異性擴增條帶。將PCR產物進行10倍梯度稀釋,設定PCR產物濃度為1,分別稀釋為1×10-1,1×10-2,1×10-3,1×10-4,1×10-5,1×10-6,1×10-7這幾個梯度濃度的DNA。幾個梯度稀釋的DNA模板以及所有GAPDH 、Ifngr1、Tlr2 、T lr4 cDNA 樣品分別配置 Realtime PCR反應體系進行PCR反應,分別在86℃、85℃、81℃、85℃收集熒光。根據繪制的梯度稀釋DNA標準曲線,各樣品目的基因和管家基因的濃度結果直接由機器生成。每個樣品的目的基因濃度除以其管家基因的濃度,即為此樣品此基因的校正后的相對含量。
2 結 果
2.1 穩定表達 Ipr1的 RAW264.7篩選 轉染pEGFP-Ipr1質粒的RAW264.7細胞經G418壓力篩選6W,獲得具有抗性的陽性克隆,經RT-PCR鑒定正確即為穩定表達Ipr1的細胞(實驗組),倒置熒光顯微鏡下觀察可見熒光集中在細胞核內見圖1-1。空質粒 pEGFP-C1轉染 RAW264.7細胞經G418篩選6W倒置熒光顯微鏡觀察可見熒光表達于整個細胞(對照組),見圖1-2。
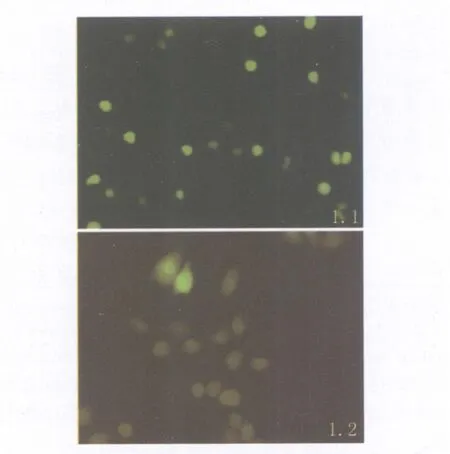
圖1.1 熒光顯微鏡觀察G418篩選6w的轉染pEGFP-Ipr1的 RAW264.7細胞 ×250圖1.2 熒光顯微鏡觀察G418篩選6w的轉染pEGFP-C1的 RAW264.7細胞 ×250Fig.1.1 6w after teansfection of RAW264.7 cellswith pEGFP-Ipr1×250Fig.1.2 6w after teansfection of RAW264.7 cellswith pEGFP-C1×250
2.2 分枝桿菌H37Ra細胞感染 實驗組細胞和對照組細胞在感染H37Ra 24h后,抗酸染色觀察可見約70%的細胞至少吞噬了1個抗酸菌,平均每個細胞吞噬20個左右的抗酸菌,表明H37Ra成功感染了實驗組和對照組RAW264.7,見圖2.1,2.2。
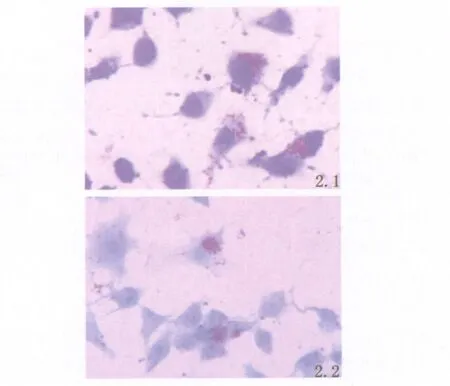
圖2.1 實驗組RAW264.7感染 H37Ra 24h;抗酸染色×1000圖2.2 對照組RAW264.7感染H37Ra 24h;抗酸染色×1000Fig.2.1 The experimental group RAW264.7 cells infected with H37Ra for 24h×1000Fig.2.2 The control group RAW264.7 cells infected with H37Ra for 24h×1000
2.3 總RNA的抽提及質量檢測 實驗組和對照組細胞抽提的總RNA,經瓊脂糖凝膠電泳觀察總RNA的28s和18s rRNA帶型清晰且28s亮度是18s亮度的2倍以上,表明總RNA較完整,見圖3。經紫外吸收測定OD值,實驗組標本和對照組標本的OD260/OD280值分別為2和1.98,表明提取的總RNA純度較高,可以用于后續實驗。實驗組標本和對照組標本總RNA濃度分別是1 765.56 ng/μ L和 1 486.74 ng/μ L。
2.4 基因芯片雜交化學發光檢測結果 實驗組標本和對照組標本所得的線性生物素標記的cRNA與小鼠Oligo Innate&Adaptive Immune Responses Microarray芯片雜交后,經化學發光檢測,X-膠片成像結果見圖4-1、4-2。

圖3 實驗組與對照組RAW264.7細胞總RNA瓊脂糖凝膠電泳結果Fig.3 Result of the experimental group cells and the control group cells total RNA
經GEArray Expression Analysis Suite軟件對原始數據進行完整的芯片數據分析,比較實驗組及對照組特定基因表達水平的差異。分析結果發現實驗組較對照組有11個基因表達上調大于2倍,其中包括T LRs及其信號通路的基因:TLR2、T LR4、Irak1、Traf6;干擾素γ受體相關基因:Ifngr1(IFN-γ R1);腫瘤壞死因子受體相關基因:Tnfrsf1a(TNFR1);細胞識別及細胞間相互作用相關基因:Lbp、Fcgr1、Colec12;炎癥反應相關的基因:Cebpe、T rem1。這些基因表達上調主要表現為調節MΦ固有免疫應答能力,而與MΦ調節T淋巴細胞的適應性免疫應答相關的一些細胞因子如 IL-1、IL-12、TNF-α等的表達卻無明顯差異。表1列出實驗組比對照組表達上調2倍以上的基因。
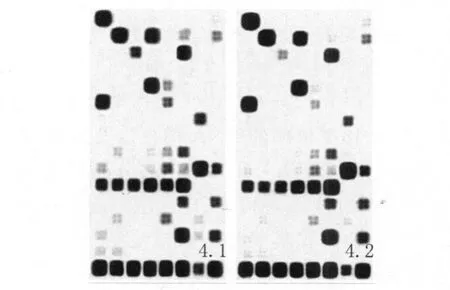
圖4-1 實驗組細胞感染H37Ra 96h后cDNA基因芯片檢測結果圖4-2 對照組細胞感染H37Ra 96h后cDNA基因芯片檢測結果Fig.4-1 cDNA expression array result of the experimental group cells infected with H37Ra for 96 hFig.4-2 cDNA expression array result of the control groupcells infected with H37Ra for 96 h
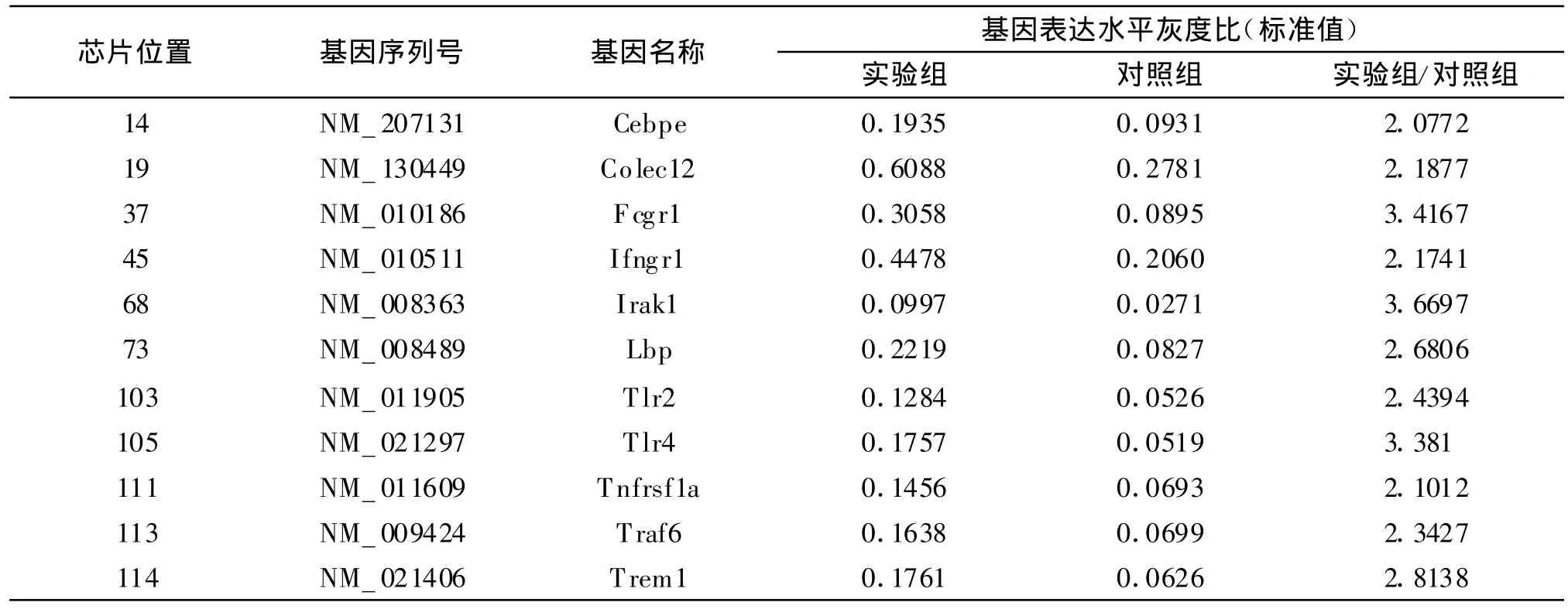
表1 實驗組比對照組表達上調2倍以上的基因Table 1 Two fold up-regulated genes between test and control
2.5 Real-time PCR檢測結果 Real-time PCR驗證芯片結果中上調的3個基因Ifngr1、Tlr2、T lr4的表達,結果用Ct值表示樣本中測定模板的起始拷貝數。Ct值越小,起始拷貝數越多,Ct值越大,起始拷貝數越小。通過制備每個目的基因的樣品標準曲線和熔解曲線保證擴增結果的準確性和專一性。標準曲線的線性回歸系數R2>0.99,擴增效率在0.83~1.0之間。結果顯示兩個樣本中內參的表達量非常接近,而Ifngr1、T lr2、Tlr4的表達量有差異,見表2。經計算其相對含量Ifngr1、Tlr2、Tlr4在實驗組的表達量分別為對照組的3.05、2.27、3.40倍,與芯片結果方向一致,結果非常接近,見表3與圖5,提示基因芯片檢測結果可靠。
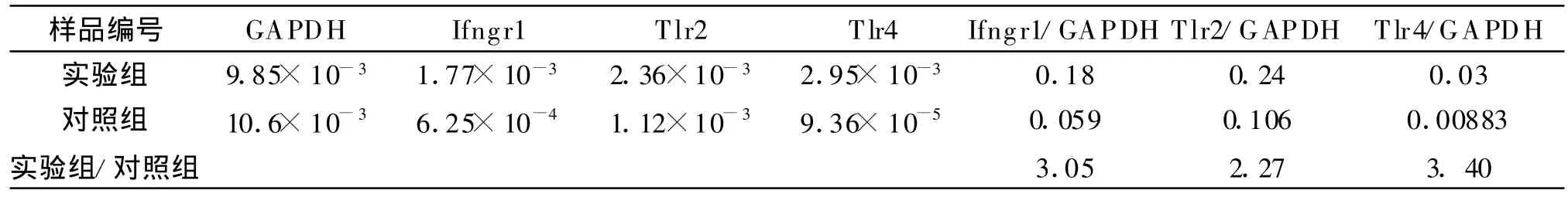
表2 Ifngr1、Tlr2、Tlr4基因在兩個樣本中的相對表達量Table 2 The relative expression level of Ifngr1 、Tlr2 、Tlr4 in the two samples
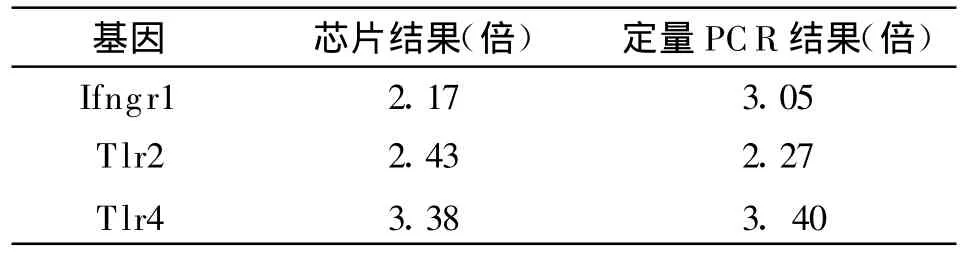
表3 Real-time PCR對基因芯片結果的驗證Table 3 Verification of the results of microarray assay by real-time PCR
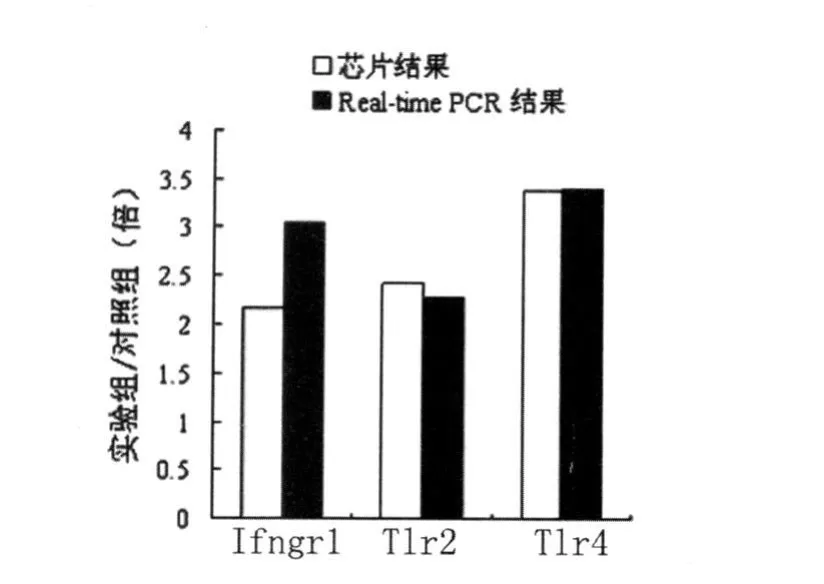
圖5 Real-time PCR對基因芯片結果的驗證Fig.5 Verification of the results of microarray assay by realtime PCR
3 討 論
Ipr1基因是新近發現的與小鼠結核病易感性相關的基因,目前的研究報道也僅限于在轉基因動物水平及體外細胞水平觀察到該基因的表達可以增強MΦ抗分枝桿菌等胞內病原體的能力,但該基因的具體作用機制還并不明確。
本實驗為了檢測Ipr1的表達對M Φ抗分枝桿菌感染相關免疫基因表達的影響及Ipr1的表達是否與M Φ抗分枝桿菌感染的已知機制間有相互關聯,我們應用了基因芯片技術。SupperArray的小鼠先天性與適應性免疫應答基因芯片(Oligo Innate&Adaptive Immune Responses Microarray)列有目前研究的已知的參與機體抗微生物感染免疫應答相關的113個基因位點,包括:IL-1R/TLRs成員及相關基因,與病原體識別有關的基因,炎癥反應相關基因,細胞凋亡相關基因,NF-κ B信號通路相關基因,細胞因子、白細胞介素、趨化因子及相應配體基因等。檢測不同標本轉錄水平免疫應答基因的表達差異。
在本實驗中,利用上述芯片檢測實驗組與對照組M Φ在感染H37Ra 96h后,相關免疫基因的表達差異。對基因芯片的結果分析我們發現Ipr1的表達主要是上調MΦ抗菌感染的免疫應答基因的表達,特別是MΦ固有免疫相關的基因表達上調尤為顯著。在表達上調的基因中,目前研究的較多的與M Φ抗Mtb等胞內病原體感染關系較密切的有:參與 TLRs信號途徑的基因 TLR2、TLR4、Irak1、Traf6;IFN-γ受體基因Ifngr1及TNF-α受體基因Tnfrsf1a。
TLR2、T LR4、Irak1、T raf6 是參與 TLRs信號途徑的基因,已經有大量的研究表明TLRs信號通路介導的MΦ抗Mtb感染在機體抗結核病的過程中發揮非常重要的作用。TLR2和T LR4是目前研究較多的與Mtb感染相關的TLRs。Mtb菌體成份或分泌成份含有一系列可以與 T LR2、T LR4相互作用的配體如LAM(脂阿拉伯甘露聚糖)、19kD脂蛋白、38kD糖脂蛋白、HSP70等,T LR2或 T LR4與相應配體結合后,可通過MyD88依賴和非依賴途徑(TLR4)激活NF-κ B,引起多種炎癥性細胞因子的釋放及上調APC細胞表面CD80、CD86等共刺激分子,在早期抗感染固有免疫殺傷病原體及晚期調節適應性免疫應答及形成肉芽腫的過程中均發揮重要作用[7]。在TLR2/T LR4介導的MyD88依賴途徑的MΦ活化過程中,M Φ內的MyD88作為一種接頭蛋白與TLR2/T LR4胞內區形成復合物,繼而招募并活化 IRAK-4、IRAK-1、T RAF-6,進一步激活下游級聯反應最終活化NF-κ B[8],最終促進M Φ活化發揮胞內殺菌及免疫調節功能。可見在此過程中 IRAK-1、T RAF-6分子的參與與活化對TLRs信號的轉導也有著重要意義。Ipr1基因上調上述基因的表達強烈提示Ipr1的表達可能與 TLR的相關基因的轉錄與表達有關,并參與TLRs介導的M Φ抗Mtb免疫應答。
γ干擾素(IFN-γ)是T細胞和NK細胞早期分泌的一種主要的Thl型細胞因子,在宿主M Φ控制Mtb感染過程中發揮非常重要的作用,IFN-γ主要通過刺激宿主MΦ產生一氧化氮合酶(NOS2)發揮抗Mtb感染作用[9]。腫瘤壞死因子α(TNF-α)是單核MΦ分泌的一種前炎癥性細胞因子,TNF-α與IFN-γ協同作用激活MΦ殺死細胞內正在復制的Mtb,并與誘導Mtb感染的M Φ凋亡密切相關[10]。本實驗發現Ipr1的表達上調了MΦIFN-γ R1基因的表達,說明Ipr1的表達還可能通過增加MΦ對IFN-γ刺激的敏感性而發揮抗菌作用,而 TNF R1基因的表達上調也提示Ipr1的表達還可能參與促進TNF-α介導的M Φ殺傷Mtb的過程及細胞凋亡。但是實驗結果中NOS2基因的表達在實驗組和對照組之間的差異卻并不明顯,也未檢測到與凋亡相關的基因 TNF-α、NF-κ B等基因的表達差異,可能是實驗設計中只是單獨應用MΦ而未加入其它刺激因素的原因。
通過本實驗檢測到了Ipr1的表達上調了一系列MΦ抗菌免疫應答基因的表達,特別值得注意的是MΦ抗微生物感染的固有免疫應答相關的基因表達,包括TLR2/T LR4基因及信號通路有關的兩個基因Irak1、Traf6,以及與MΦ活化及殺傷胞內病原體有關的細胞因子受體:Ifngr1、Tnfrsf1a。而參與MΦ調節適應性免疫應答的細胞因子有關基因IL-1、TNF-α、IL-12等表達無明顯差異。提示 Ipr1的表達可能主要是通過增強MΦ固有免疫應答的殺傷吞噬的Mtb機制,尤其是TLRs活化機制的作用進而增強機體抗Mtb感染的能力,但究竟Ipr1是如何與T LRs活化機制間發生相互作用的以及Ipr1表達產物的作用位點仍需進一步的研究。
[1]WHO.Global tuberculosis control:WHO report 2010.
[2]Sundaramurthy V,Pieters J.Interactions of pathogenic mycobacteria with host macrophages[J].Microbes Infect,2007,9(14-15):1671-1679.
[3]Fremond CM,Yeremeev V,Nicolle DM,et al.FatalMycobacterium tuberculosisinfection despite adaptive immune response in the absence of MyD88[J].J Clin Invest,2004,114(12):405-412.
[4]Pan H,Yan BS,Rojas M,et al.gene mediates innate immunity to Tuberculosis[J].Nature,2005,434(7034):767-772.
[5]李娜,朱道銀,帖儒修,等.小鼠Ipr1基因 EGFP基因融合表達載體的構建及在鼠巨噬細胞中的表達[J].細胞與分子免疫學雜志,2008,24(3):231-233.
[6]師長宏,張海,席麗,等.結核分枝桿菌 Hsp65與IL-2融合蛋白穩定表達細胞系的建立[J].中國人獸共患病學報,2008,24(8):718-721.
[7]Quesniaux V,Fremond C,Jacobs M,et al.T oll-like receptor pathways in the immune responses to mycobacteria[J].Microbes Infect.2004,6(10):946-959.
[8]Zhu J,M ohan C.Toll-like receptor signaling pathways-therapeutic opportunities[J].J Immunol,2010,2010:7812-35.
[9]Feng CG,Kaviratne M,Rothfuchs AG,et al.NK cell-derived IFN-gamma differentially regulates innate resistance and neutrophil response in T cell-deficient hosts infected with Mycobacterium tuberculosis[J].J Immunol,2006,77(10):7086-7093.
[10]Arcila M L,S nchez MD,O rtiz B,et al.Activation of apoptosis,but not necrosis,duringMy cobacterium tuberculosisinfection correlated with decreased bacterial g rowth:role of TNF-alpha,IL-10,caspases and phospholipase A2[J].Cell Immunol,2007,249(2):80-93.
Immune response genes expressing of Ipr1 expressed macrophage anti-M.tuberculosis
LI Na,LIU Peng-fei,LI Bo-qing,QIAO Yuan-yuan,ZHANG Yu-mei
(Department of Pathogeny Biology of Binzhou Medical College,Yantai264003,China)
To study the effect and mechanisms of Ipr1 gene expressing in macrophages infected withMycobacterium tuberculosisH37Ra use cDNA microarry.The experimental group RAW264.7 cells were transfected with pEGFP-Ipr1 and the stable Ipr1 gene expressed RAW264.7 cells were selected by G418.The control group RAW264.7 cells were transfected with pEGFP-C1and selected by G418.Experimental group and control group cells were infected byMycobacterium tuberculosisH37Ra.After 96h infection,the total RNA were isolated from the two groups.Then using a cDNA microarry(Mouse Innate and Adaptive Immune Responses Microarray113 genes)to detect the differences of experimental group and control group in gene expression related to immune response.Real-time quantitative PCR test was used to verify the reliability of the chip results.The cDNA microarry result display that Ipr1 gene expression up regulated 11 genes for macrophage anti-Mtbinnate immunity involved TLRs signaling pathway:TLR2 and T LR4,Irak1,T raf6;as well as interferon-related genes:Ifngr1(IFN-γ R1)and tumor necrosis factor-related genes:Tnfrsf1a(TNFR1),et al.And there was no significant difference about the molecular expression involved in the regulation of the adaptive immune response such as:IL-1,IL-12.The results of real-time PCR reaction TLR2 and TLR4 and Ifngr1 indicated that the real-time PCR results consistent with the trend of chip results.The results of gene chip analysis point that the Ipr1 gene expression maybe enhance macrophage anti-Mtbinfection by the mechanisms of increasing the innate immunity gene expression,in particular TLR2/TLR4 and its signal transduction molecules as well as the IFN-γ R1,TNFR1 expression,and promote macrophage activation and intracellular anti-Mtbeffect.This constitutes the basis for further study of the mechanisms of Ipr1 gene in host innate immunity against tuberculosis infection.
Ipr1;Mycobacterium tuberculosis;macrophage
R378.91
A
1002-2694(2011)08-0715-06
*山東省高等學校科技計劃資助項目(J09LF01)
1.濱州醫學院病原生物學教研室,煙臺 264003;Email:bzy xy lina@163.com
2.濱州醫學院附屬醫院神經外科,煙臺 264003
2011-03-09;
2011-05-21
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
