瘦素對L-02肝細胞甘油三酯沉積及乙酰輔酶A羧化酶表達的影響
2011-06-18 03:45:56周紅宇陽學風
實用藥物與臨床 2011年5期
周紅宇,陽學風
非酒精性脂肪肝(NAFLD)是一種肝組織學改變的臨床病理綜合征,與酒精性肝病相類似,但與飲酒無明確關系。根據其病理學改變及臨床表現,分為單純性脂肪肝、脂肪性肝炎、脂肪性肝纖維化、脂肪性肝硬化[1]。肝臟所含脂質絕大部分為甘油三酯,各種致病因素均可導致肝細胞內甘油三酯(TG)異常堆積,從而導致FLD的發生。
瘦素是肥胖基因(Obese gene)編碼的蛋白產物,受體廣泛分布于脂肪、骨骼肌、肝臟、心、肺、胰、卵巢、睪丸等組織。瘦素通過與受體結合發揮作用[2],調節機體脂肪的穩定,抑制食欲、增加能耗和減少體重可能是瘦素的主要作用。本文以人肝L-02脂肪變細胞模型,通過油紅“O”染色、RTPCR等方法,對瘦素作用后的脂肪變細胞進行研究,了解瘦素與NAFLD的關系,并初步探討瘦素對其的治療作用。
1 實驗方法
1.1 非酒精性脂肪變肝細胞模型的建立 人肝L-02細胞株培養接種于50mL培養瓶中,在含體積分數10%的滅活胎牛血清、100 U/mL青霉素和100μg/mL鏈霉素的 1640完全培養基中,于37℃,5%CO2培養,飽和濕度,貼壁生長。待細胞長滿80%瓶底,消化液消化傳代。在更換新鮮培養液后加入胎牛血清,使其濃度為50%,繼續培養24 h,形成非酒精性肝細胞脂肪變性模型。
1.2 造模是否成功的判斷 Lillie氏油紅-O染色:將細胞培養于有無菌蓋玻片的6孔培養板,用PBS沖洗蓋玻片5min(×3次),50%異丙醇固定1min,油紅“O”染色10min,蒸餾水沖洗1min(×3次),蘇木素染色5min,分色和返藍后,HPIAS-1000型圖像分析系統收集圖像。
1.3 RT-PCR法檢測人肝L-02乙酰輔酶A羧化酶(ACC)mRNA的表達 以 lμg逆轉錄產物(cDNA)為模板,通過半定量RT-PCR,以三磷酸甘油醛脫氫酶(GAPDH)作為內參照。
2 結果
2.1 人肝L-02細胞脂肪化 正常細胞(圖1)用50%胎牛血清培養基培養24 h后,用油紅“O”染色,顯微鏡下觀察,細胞漿內有大量脂滴存在,符合脂肪化肝細胞特點(圖2)。

圖1 正常肝細胞(油紅‘O’×400)
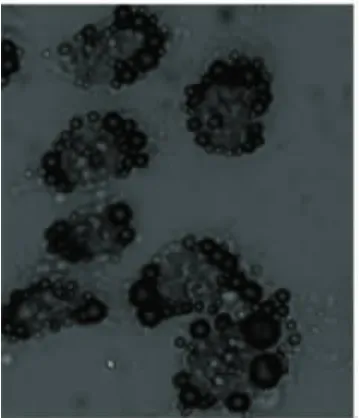
圖2 模型組(油紅‘O’×400)
2.2 不同瘦素劑量干預后脂肪化人肝L-02細胞的變化 脂肪化人肝L-02細胞分別用10-7mol/L、10-6mol/L、10-5mol/L 瘦素 +1640 完全培養基培養24 h后,用油紅“O”染色,顯微鏡下觀察,脂肪化程度逐漸減輕,見圖3~圖5。
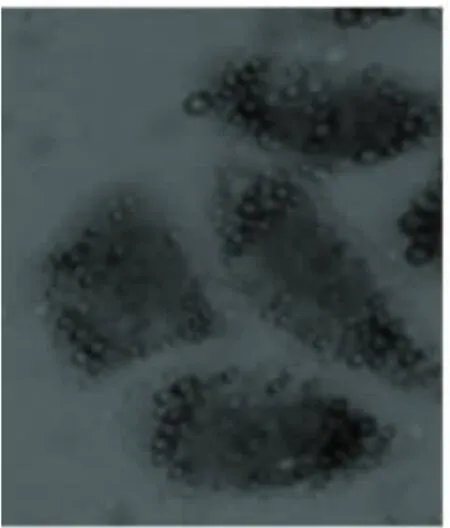
圖3 模型組+瘦素10-7mol/L組(油紅‘O’×400)

圖4 模型組+瘦素10-6mol/L組
2.3 瘦素對人肝L-02脂肪變細胞乙酰輔酶A羧化酶mRNA表達的影響 用RT-PCR檢測人肝L-02細胞乙酰輔酶A羧化酶mRNA的表達。結果顯示,經瘦素處理的人肝L-02細胞乙酰輔酶A羧化酶mRNA的表達與瘦素呈濃度依賴性遞減。模型組、10-7mol/L、10-5mol/L 及10-5mol/L 瘦素處理組目的基因及GAPDH基因灰度值比值分別為0.867、0.745、0.642、0.512,正常組及正常細胞加瘦素組分別為0.460及0.437,見圖6。瘦素處理組與模型組相比,差異有統計學意義(P<0.05)。
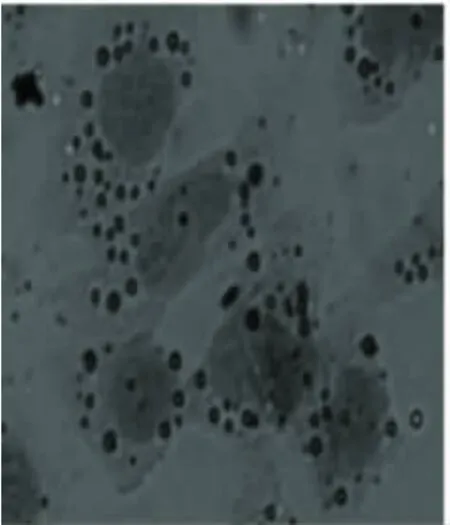
圖5 模型組+瘦素10-5mol/L組
圖6 不同濃度瘦素處理對人肝L-02脂肪變細胞ACCmRNA表達的影響(灰度值)
3 討論
近年來,隨著生活水平的提高,飲食結構的變化,檢測手段的提高,非酒精性脂肪肝(NAFLD)發病率呈逐年上升趨勢。自1994年起,一系列研究結果表明,NAFLD的發病無性別差異,在非肥胖和糖尿病人群中也有較高的發病率。特別是發現NAFLD可進展至肝炎、肝纖維化和肝硬化之后[3-4],逐漸為人們所重視。約75%的2型糖尿病患者可發生不同形式的NAFLD;70%的肥胖患者發生脂肪肝,其中18%出現 NASH[5]。NAFLD的發病機制尚不十分清楚,人們廣泛認同Day等[6]提出的“二次打擊”理論。多因素聯合作用誘發胰島素抵抗(IR),后者通過增加脂肪分解和FFAs轉運至肝臟導致第一次打擊—脂肪肝形成,并造成肝臟對第二次打擊的敏感性增強。氧應激、脂質過氧化等進一步促進IR,加重氧應激和肝細胞損傷,導致炎癥反應、肝細胞變性和肝纖維化,即第二次打擊。本研究結合文獻方法[7],用高脂培養基造模,形成典型的非酒精性脂肪肝細胞模型,主要研究NAFLD形成的第一步肝細胞內脂肪沉積及瘦素干預后的變化。
肝臟是體內參與脂質代謝的重要場所。肝臟脂質代謝穩態的變化是構成各種形式脂肪肝的基礎,各種致病因素可通過一個或多個環節導致肝細胞內TG異常堆積。肝細胞不能儲存脂肪,TG在肝細胞內,一方面氧化分解供能,另一方面與載脂蛋白B100、載脂蛋白C等以及磷脂、膽固醇結合生成極低密度脂蛋白(VLDL),由肝細胞分泌通過血液循環運輸至肝外組織。當肝細胞合成TG增多,TG轉運或氧化障礙時,均可導致TG的堆積,形成脂肪肝。
本實驗發現肝細胞普遍發生脂肪變性,細胞腫脹,內含較大的脂肪滴,可將細胞核擠壓向胞膜,表明高脂環境可引起肝臟脂肪浸潤(TG貯存)。而經過瘦素處理后,肝細胞內脂滴明顯減少,細胞形態恢復正常,與瘦素呈劑量依賴性關系。說明10-7~10-5mol/L濃度的瘦素對離體脂肪變肝細胞有積極的防治作用。
瘦素是一種主要由白色脂肪組織分泌合成、由167個氨基酸殘基組成的蛋白質類激素[8]。主要在白色脂肪組織中表達,通過靶細胞膜上的受體及相應的信號轉導體系發揮作用。OB-Rb主要存在于下丘腦中能表達NPY的細胞表面,與瘦素結合后,通過Janus激酶(JAK)以及信號轉導和轉錄激活物(STAT)蛋白發揮信號轉導作用,調控核內DNA上leptin效應基因的轉錄活性。瘦素與特異性運輸蛋白結合,通過血腦屏障后與下丘腦特異受體結合,把體脂信號傳遞給下丘腦。下丘腦再通過一系列神經體液因素調節機體能量代謝,調整體脂,控制體重相對恒定[9]。給OB小鼠腦脊液中注入瘦素后[10],進食明顯減少,體重逐漸下降,減少的僅為脂肪組織,且基礎代謝率有所升高,說明抑制食欲、增加能耗和減少體重可能是瘦素減肥的主要機制。
瘦素可能從外周直接拮抗肝脂肪變性。瘦素可以激活AMP活化蛋白激酶,減少脂肪沉積[11],還可特異性抑制肝臟的硬脂酰基-輔酶A去飽和酶,使肝臟 VLDL生成和脂肪沉積減少[12]。Unger等[13]認為,非脂細胞瘦素受體發生功能障礙時,細胞內TG含量能增加100倍,而持續表達的異位高瘦素血癥能使細胞內TG耗竭;瘦素還干預胰島素在肝臟中的作用,一方面拮抗胰島素誘導的PEPCK下調,抑制胰島素受體底物-1(IRS-1)的磷酸化,影響胰島素受體后信號轉導;另一方面,瘦素通過對PEPCK及糖異生的影響,限制TG的合成,提高肝臟及外周組織對胰島素的敏感性。Stumvoil等[14]發現,瘦素與脂聯素等連用,能促進小鼠肝脂肪燃燒及能量消耗,可逆轉缺乏過氧化物酶體增殖受體(PPAR)α的脂肪萎縮小鼠的胰島素抵抗。
本實驗證實細胞內TG明顯增加,經過瘦素處理后,肝細胞內TG明顯減少,與瘦素呈劑量依賴性關系。說明瘦素能明顯減少離體脂肪變肝細胞的TG沉積,與文獻[12-15]報道一致。本實驗顯示,瘦素對離體脂肪變肝細胞具有降低細胞內脂肪沉積的作用,對TG的作用明顯。同時,本實驗通過油紅“O”染色顯示,瘦素處理后脂肪變肝細胞內TG量明顯減少,瘦素可抑制乙酰CoA羧化酶的表達而抑制TG的合成,可見瘦素可使離體肝細胞內TG合成減少,減少TG沉積,從而減輕肝細胞脂肪化程度。但其對人類非酒精性脂肪性肝臟疾病是否具有治療意義,以及具體作用機制,有待進一步研究。
[1]周峘,滕香宇,劉偉.2型糖尿病和非酒精性脂肪肝[J].中國臨床醫學,2007,14(5):675-677.
[2]Day CP.Non-alcoholic steatohepatitis(NASH):where are we now and where are we going[J].Gut,2002,50(5):585-588.
[3]郭宏,許傳芬,郭冬.非酒精性脂肪肝患者肝損害程度與代謝綜合征各組分的關系[J].中國醫藥,2010,5(12):1139-1141.
[4]楊秉輝.非酒精性脂肪性肝病與肝癌[J].中國臨床醫學,2011,18(1):1-2.
[5]Medina J,Fernández-Salazar LI,García-Buey L,et al.Approach to the pathogenesis and treatment of nonalcoholic steatohepatitis[J].Diabetes Care,2004,27(8):2057-2066.
[6]Day CP,James OF.Steatohepatitis:a tale of two“hits”[J].Gastroenterology,1998,114(4):842-845.
[7]徐正捷,范建高,王國良,等.乳果糖對大鼠非酒精性脂肪性肝炎模型形成的影響[J].中華消化雜志,2003,23:97-100.
[8]臧謀圣,徐勝前.瘦素和瘦素受體在類風濕性關節炎及其骨質疏松發病機制中的研究進展[J].中國醫藥,2010,5(3):281-283.
[9]鄭素梅,梁麗霞,鄭風穎,等.瘦素與骨代謝[J].中國醫藥,2009,4(3):238-239.
[10]Le Lay S,Boucher J,Rey A,et al.Decreased resistin expression in mice with different sensitivities to a high-fat diet[J].Biochem Biophys Res Commun,2001,289[2]:564-567.
[11]Minokoshi Y,Kim YB,Peroni OD,et al.Leptin stimulates fatty-acid oxidation by activating AMP-activated protein kinase[J].Nature,2002,415[6869]:339-343.
[12]Oral EA,Simha V,Ruiz E,et al.Leptin-replacement therapy for lipodystrophy[J].N Engl J Med,2002,346:570-578.
[13]Uygun A,Kadayifci A,Yesilova Z,et al.Serum leptin levels in patients with nonalcoholic teatohepatitis[J].Am J Gastraenterol,2000,95(12):3584-3589.
[14]Stumvoil M,Tschritter O,et al.Association of T-G polymorphism in adiponection(exon 2)with obesity and insulin sensitivity:interaction with family history of type 2 diabetes[J].Diabetes,2002,51(1):37-41.
[15]張霞意,郭家偉,羅紅彬,等.非酒精性脂肪肝與血清瘦素和可溶性瘦素受體的關系[J].實用醫學雜志,2009,25(17):2849-2851.
