匹諾塞林對體外共培養(yǎng)海兔SN/L7神經(jīng)元電生理活動的作用
2011-06-09 02:10:54胡江原SamuelSchacher杜冠華
中國藥理學(xué)通報(bào) 2011年6期
應(yīng) 劍,胡江原,陳 陽,Samuel Schacher,杜冠華
(1.中國醫(yī)學(xué)科學(xué)院藥物研究所國家藥物篩選中心,北京 100050;2.美國哥倫比亞大學(xué)醫(yī)學(xué)中心神經(jīng)科學(xué)系,紐約 10032)
海洋軟體動物海兔(aplysia californica)神經(jīng)系統(tǒng)簡單,神經(jīng)細(xì)胞胞體大且數(shù)量少、便于實(shí)驗(yàn)操作,是研究學(xué)習(xí)記憶和神經(jīng)功能的重要模式生物。20世紀(jì)60年代,Kandel等[1]對海兔縮鰓反射的神經(jīng)通路進(jìn)行研究,發(fā)現(xiàn)來自噴水管的感覺信息經(jīng)胸膜神經(jīng)節(jié)的感覺神經(jīng)元SN傳至腹部神經(jīng)節(jié)的L7運(yùn)動神經(jīng)元,L7支配鰓肌,控制縮鰓運(yùn)動。80年代,Glanzman等[2]建立了SN和L7的共培養(yǎng)模型(SN/L7),即分別分離L7和單個SN后,轉(zhuǎn)移至玻璃底培養(yǎng)皿中共同培養(yǎng),用玻璃微電極將SN和L7的軸突輕輕搭在一起,在適當(dāng)?shù)呐囵B(yǎng)條件下,SN和L7之間可形成特異的突觸連接。
介導(dǎo)SN/L7突觸瞬時興奮傳遞的遞質(zhì)是谷氨酸[3]。谷氨酸作用于突觸后膜的興奮性氨基酸受體,使突觸后膜對Na+、K+的通透性增加,Na+內(nèi)流大于K+外流,突觸后膜局部去極化,從而產(chǎn)生興奮性突觸后電位(EPSP)。在適當(dāng)條件下,SN/L7可表現(xiàn)出短時程或者長時程的突觸可塑性,模擬自然條件下海兔縮鰓反射的習(xí)慣化和敏感化。短時程突觸可塑性主要依賴于離子通道的修飾和遞質(zhì)釋放的變化,通過一系列下游信號通路的轉(zhuǎn)導(dǎo),還可能對長時程突觸可塑性產(chǎn)生影響。
匹諾塞林(PNCB)是蜂膠中含量最高的黃酮類化合物[4],具有抗炎[5]、殺菌[6-7]、抗氧化[8]、舒張血管[9]、神經(jīng)保護(hù)[10-12]等廣泛的藥理作用。在藥物篩選以及對PNCB抗腦缺血損傷機(jī)制的研究中,發(fā)現(xiàn)PNCB作用于K+通道、Ca2+通道、NMDA受體、GABA受體等多種離子通道。本實(shí)驗(yàn)通過考察PNCB對海兔SN/L7的電生理活動的作用,探討PNCB對海兔神經(jīng)細(xì)胞短時程突觸可塑性和長時程突觸可塑性的影響,為PNCB對神經(jīng)系統(tǒng)作用機(jī)制的研究提供參考依據(jù)。
1 材料與方法
1.1材料不同年齡段的加州海兔,分別用于分離L7細(xì)胞(幼年海兔,體質(zhì)量2 g)、SN細(xì)胞(成年海兔,體質(zhì)量60~80 g),抽取淋巴血液(成年海兔,體質(zhì)量約1 kg),均購于美國邁阿密大學(xué)海兔國家資源庫。實(shí)驗(yàn)中所用試劑均購于美國Sigma公司。PNCB(純度大于98%)由中國醫(yī)學(xué)科學(xué)院藥物研究所吳松教授提供。
1.2SN/L7細(xì)胞培養(yǎng)根據(jù)SN/L7共培養(yǎng)的實(shí)驗(yàn)方法[2],從成年海兔的胸膜神經(jīng)節(jié)分離SN,從幼年海兔的腹部神經(jīng)節(jié)分離L7。將兩個神經(jīng)細(xì)胞移至同一個玻璃底培養(yǎng)皿中,用微電極輕輕撥動軸突,使兩個神經(jīng)細(xì)胞的軸突相互接觸,并于18℃靜置培養(yǎng)。細(xì)胞培養(yǎng)基為含50%海兔淋巴血液和100 mmol·L-1L-谷氨酸的L15培養(yǎng)基。共培養(yǎng)體系隔天換液,d 5開始實(shí)驗(yàn)。
1.3電生理技術(shù)應(yīng)用全細(xì)胞電生理技術(shù)記錄SN/L7的興奮性突觸后電位(EPSP)[13]。將刺激電極靠近SN的細(xì)胞體,設(shè)定工作電位為-85 mV,對SN胞體施加電刺激,每個刺激持續(xù)0.3~0.5 ms;將記錄電極插入L7的細(xì)胞體引出EPSP。
1.4匹諾塞林對SN/L7的電生理活動的影響將50%的L15培養(yǎng)基與50%的人工海水(ASW)混合均勻,即 L15/ASW 工作液。精密稱取 PNCB,用DMSO 梯度稀釋至 10、20、30、40、100、200、300、400 mmol·L-1,使用前再用 L15/ASW 稀釋1 000倍待用。細(xì)胞培養(yǎng)至d 5時,用L15/ASW輕輕洗去培養(yǎng)基,記錄SN/L7的初始EPSP;然后用10ml含不同濃度PNCB的L15/ASW溶液替換L15/ASW,室溫靜置5 min后,再記錄EPSP。對于PNCB給藥濃度為0.1~0.4 mmol·L-1的實(shí)驗(yàn)組,用L15/ASW充分洗去藥物,換成細(xì)胞培養(yǎng)基,放回培養(yǎng)箱培養(yǎng),并分別于 1、24 h 記錄 EPSP。另取 0.01 mmol·L-1PNCB孵育過的SN/L7,洗去或不洗去藥物,加入0.005 mmol·L-15-HT 溶液孵育5 min,記錄 EPSP。
另取一批細(xì)胞,于d 5用L15/ASW洗去細(xì)胞培養(yǎng)基后,記錄 SN/L7的初始 EPSP。再用含0.01 mmol·L-1PNCB 的 L15/ASW 孵育 SN/L7,每 20 min換液1次,共5次。然后用L15/ASW洗去藥物,換回細(xì)胞培養(yǎng)基于18℃繼續(xù)培養(yǎng)。d 6重復(fù)以上孵育步驟。d 7用L15/ASW洗去細(xì)胞培養(yǎng)基后,記錄EPSP。
2 結(jié)果
2.1匹諾塞林降低SN/L7的興奮性突觸后電位(EPSP)的作用SN和L7轉(zhuǎn)移至玻璃底培養(yǎng)皿后,開始貼壁生長,搭在一起的軸突之間逐漸形成突觸。伴隨著突觸成熟,可以記錄到幅值越來越高的EPSP。d 4~5,細(xì)胞成熟(Fig 1),EPSP的幅值也基本穩(wěn)定[14]。
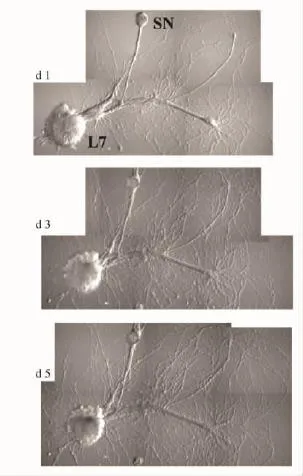
Fig 1 Typical SN/L7 co-culture at day 1,3 and 5
使用培養(yǎng)至d 5的細(xì)胞,考察不同濃度的PNCB對SN/L7的EPSP的影響。以PNCB作用前細(xì)胞的初始EPSP幅值為基準(zhǔn)值,計(jì)算藥物作用后的EPSP幅值相對于初始值的百分比。對照組細(xì)胞用含0.1%的DMSO的L15/ASW孵育5 min后,用同樣的指標(biāo)計(jì)算EPSP幅值的變化。在檢測時間內(nèi),對照組的EPSP幅值略有增高,但無顯著性變化;而PNCB(0.01 ~0.4 mmol·L-1)作用5 min,使 SN/L7的峰電位消失、突觸電位降低(Fig 2)。
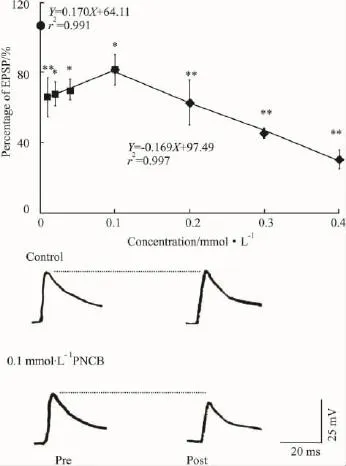
Fig 2 Normalized EPSP amplitude(%pretreatment baseline)in Aplysia SN/L7 co-cultures after 5 min incubation of 0.01 ~ 0.4 mmol·L-1pinocembrin
當(dāng) PNCB濃度低于0.1 mmol·L-1時,其降低EPSP的作用強(qiáng)度與給藥劑量呈負(fù)相關(guān)的線性關(guān)系(r>0.995)。而在 0.1 ~0.4 mmol·L-1的濃度范圍內(nèi),PNCB降低EPSP的作用強(qiáng)度與給藥劑量呈正相關(guān)的線性關(guān)系(r>0.998)。當(dāng)PNCB濃度為0.4 mmol·L-1時,EPSP的幅值僅為初始幅值的(30.7±5.1)%(P<0.01)。此外,當(dāng)藥物濃度 >0.1 mmol·L-1時,細(xì)胞靜息電位也受到影響,觀察到一定程度的去極化。
2.2匹諾塞林可逆性降低EPSP幅值的作用PNCB與海兔神經(jīng)細(xì)胞作用后,洗去含 PNCB的L15/ASW,置換成細(xì)胞培養(yǎng)基,于18℃繼續(xù)培養(yǎng)1 h,EPSP的幅值即回升為初始幅值的71% ~89%;繼續(xù)培養(yǎng)24 h,則大多數(shù)細(xì)胞的EPSP幅值恢復(fù)至初始值。結(jié)果表明,PNCB對海兔神經(jīng)細(xì)胞EPSP幅值的抑制作用是可逆的,將PNCB洗去后EPSP可以得到恢復(fù)。而動作電位幅度的恢復(fù)與PNCB的濃度無直接關(guān)系(Tab 1)。
2.3匹諾塞林抑制SN/L7對5-HT的反應(yīng)性的作用海兔神經(jīng)系統(tǒng)通過一個中間神經(jīng)元釋放5-HT至SN和L7的突觸連接,易化突觸傳遞。在體外培養(yǎng)體系中,用0.005 mmol·L-1的5-HT刺激細(xì)胞,可以使細(xì)胞的EPSP升高,甚至出現(xiàn)尖銳的峰電位(Fig 3A)。用 0.02 mmol·L-1PNCB 孵育細(xì)胞5min使SN/L7的EPSP幅值下降,再加入5-HT(終濃度為0.005 mmol·L-1)孵育5 min后,未見EPSP幅值升高或峰電位出現(xiàn)(Fig 3B)。撤除PNCB后靜置5 min再給予5-HT刺激,則可以記錄到EPSP幅值增加或者峰電位(Fig 3C)。
Tab 1 Normalized EPSP amplitude in Aplysia SN/L7 co-cultures after removal of pinocembrin(±s,n=3)

Tab 1 Normalized EPSP amplitude in Aplysia SN/L7 co-cultures after removal of pinocembrin(±s,n=3)
The initial EPSP level of each culture is marked as 100%
Pinocembrin concentration/mmol·L -1 Normalized EPSP amplitude/%0 1 h 24 h 0.1 81.4 ±8.6 77.7 ±23.6 96.7 ±4.7 0.2 62.8 ±12.8 71.5 ±18.2 87.3 ±5.1 0.3 45.4 ±2.6 71.6 ±3.8 106.3 ±0.1 0.4 30.7 ±5.1 88.8 ±9.8 105.8 ±3.0

Fig 3 Pinocembrin inhibited the spike evoked by 5-HT in Aplysia SN/L7 co-cultures.
2.4匹諾塞林對SN/L7長時程突觸可塑性的作用
為考察PNCB是否影響SN/L7的長時程突觸可塑性,在d 5和d 6分別在細(xì)胞培養(yǎng)基中加入0.01 mmol·L-1PNCB,并在 d 6 給藥24 h 后記錄 EPSP。結(jié)果表明,對照組的EPSP幅值為給藥前的(104.1±10.5)%;PNCB給藥組的EPSP幅值為給藥前的(96.6±4.7)%。兩個實(shí)驗(yàn)組EPSP的變化差異沒有顯著性。
3 討論
負(fù)責(zé)海兔SN/L7突觸興奮性傳遞的主要遞質(zhì)是谷氨酸。海兔神經(jīng)細(xì)胞的突觸后膜上存在NMDA、AMPA型等谷氨酸受體[15]。本實(shí)驗(yàn)證明,給予5 min 0.01~0.4 mmol·L-1PNCB 的刺激,可抑制海兔SN/L7的電生理效應(yīng),使EPSP幅值降低。這一現(xiàn)象表明,PNCB對SN/L7的興奮性突觸傳遞有一定程度的抑制作用,其抑制SN/L7突觸的興奮性的作用可能與NMDA型谷氨酸受體有關(guān)。
以0.1 mmol·L-1為轉(zhuǎn)折點(diǎn),抑制作用呈現(xiàn)兩相性,即PNCB 的濃度在0.01~0.1 mmol·L-1的范圍內(nèi),抑制作用隨著藥物濃度增加而減弱;而當(dāng)PNCB的濃度在0.1~0.4 mmol·L-1的范圍內(nèi)時,抑制效應(yīng)隨著藥物濃度增加而增強(qiáng)。且這兩相均呈現(xiàn)良好的線性相關(guān)性。因此,PNCB抑制SN/L7興奮性突觸傳遞的作用在不同濃度下可能有不同的機(jī)制,需要進(jìn)一步研究。近幾年的研究發(fā)現(xiàn),神經(jīng)細(xì)胞NMDA受體可分為突觸型(NMDARs)與非突觸型(NMDARn)兩類。NMDA抑制劑,如美金剛等,在低濃度時優(yōu)先抑制NMDARn,對NMDARs的抑制較弱,使得NMDARs相對活化;隨著抑制劑的濃度增加,NMDARs才被明顯抑制[16]。這一現(xiàn)象有助于解釋本實(shí)驗(yàn)中觀察到的PNCB抑制海兔SN/L7 EPSP的兩相性。
神經(jīng)細(xì)胞SN/L7與PNCB作用后,撤除PNCB可以使神經(jīng)細(xì)胞的EPSP幅值恢復(fù)至初始值。這一結(jié)果提示,PNCB對其作用靶點(diǎn)的抑制作用是可逆的。
5-HT有易化SN/L7神經(jīng)突觸的作用。生理狀態(tài)下,用0.005 mmol·L-15-HT孵育SN/L7共培養(yǎng)體系5min,即可記錄到EPSP增強(qiáng)。PNCB的作用使SN/L7對5-HT的反應(yīng)性消失;而撤除PNCB后,這一反應(yīng)性迅速得以恢復(fù),提示PNCB對5-HT易化神經(jīng)突觸的通路也有可逆的抑制作用。用5-HT刺激體外SN/L7共培養(yǎng)細(xì)胞,模擬的是海兔縮鰓反射的敏感化。5-HT作用于SN/L7的突觸前膜,激活腺苷酸環(huán)化酶,繼而使蛋白激酶磷酸化,磷酸化的蛋白激酶作用于突觸前膜的鉀離子通道使之關(guān)閉,鉀離子外流減少,使動作電位到來之前的去極化時間延長,從突觸前膜釋放的谷氨酸增加,EPSP的幅值隨之增加,突觸效能增強(qiáng)[17-18]。因此,PNCB 可逆地抑制SN/L7共培養(yǎng)體系對5-HT的反應(yīng)性,可能與PNCB對突觸后膜谷氨酸受體的可逆性抑制有關(guān);此外,由于PNCB具有開放鉀離子通道的作用[9],PNCB也可能通過開放突觸前膜的鉀離子通道,減少谷氨酸釋放,實(shí)現(xiàn)其抑制作用。
模擬5-HT誘導(dǎo)長時程易化的實(shí)驗(yàn)方法,連續(xù)兩天用0.01 mmol·L-1PNCB孵育SN/L7細(xì)胞,24 h后 SN/L7的 EPSP幅值沒有明顯變化。表明PNCB對NMDA受體的抑制作用未在基因和蛋白水平產(chǎn)生更深遠(yuǎn)的影響。
此外,實(shí)驗(yàn)中還觀察到高濃度PNCB影響L7靜息電位,表現(xiàn)為部分去極化,表明高濃度PNCB可能在突觸后膜影響鉀離子外流或鈉離子內(nèi)流。由于有研究證明PNCB可開放鉀離子通道,因此,PNCB可能通過部分開放L7細(xì)胞膜上的鈉離子通道,影響其靜息電位。
綜上所述,PNCB作用于其SN/L7模型,總體表現(xiàn)為對突觸興奮性傳導(dǎo)的可逆性抑制。但是,在實(shí)驗(yàn)劑量下,PNCB不影響SN/L7的長時程突觸可塑性。PNCB是一種藥理作用廣泛的化合物。本實(shí)驗(yàn)室在研究PNCB作用機(jī)制的工作中,發(fā)現(xiàn)PNCB與一系列病理通路關(guān)聯(lián)。其中,PNCB抗腦缺血損傷的作用與NMDA受體相關(guān)。但是,PNCB藥理作用的靶點(diǎn)究竟何在,仍然未有定論。本研究提示,PNCB可能通過作用于突觸前膜和突觸后膜的多種離子通道,產(chǎn)生復(fù)雜的瞬時效應(yīng)以及下游效應(yīng)。下一步工作將深入探討PNCB對離子通道的作用,相信不日將尋得PNCB的作用靶點(diǎn)。
[1]Kupfermann I,Pinsker H,Castellucci V,Kandel E R.Central and peripheral control of gill movements in aplysia[J].Science,1971,174(15):1252-6.
[2]Glanzman D L,Kandel E R,Schacher S.Identified target motor neuron regulates neurite outgrowth and synapse formation of aplysia sensory neuronsin vitro[J].Neuron,1989,3(4):441 -50.
[3]Dale N,Kandel E R.L-glutamate may be the fast excitatory transmitter of Aplysia sensory neurons[J].Proc Natl Acad Sci USA,1993,90(15):7163-7.
[4]Bankova V S,Popov S S,Marekov N L.High-performance liquid chromatographic analysis of flavonoids from propolis[J].J Chromatogra A,1982,242(1):135-43.
[5]Sala A,Recio M C,Schinella G R,et al.Assessment of the antiinflammatory activity and free radical scavenger activity of tiliroside[J].Eur J Pharmacol,2003,461(1):53 -61.
[6]Pepeljnjak S,Jalsenjak I,Maysinger D.Flavonoid content in propolis extracts and growth inhibition of bacillus subtilis[J].Pharmazie,1985,40(2):122 -3.
[7]Del Rayo C M,Sanchez B,Quiroz H,et al.Pinocembrine:a bioactive flavanone from teloxys graveolens[J].J Ethnopharmacol,1991,31(3):383-9.
[8]Santos A C,Uyemura S A,Lopes J L,et al.Effect of naturally occurring flavonoids on lipid peroxidation and membrane permeability transition in mitochondria[J].Free Radic Biol Med,1998,24(9):1455-61.
[9]Zhu X M,F(xiàn)ang L H,Li Y J,Du G H.Endothelium-dependent and-independent relaxation induced by pinocembrin in rat aortic rings[J].Vascul Pharmacol,2007,46(3):160 -5.
[10]Liu R,Gao M,Yang Z H,Du G H.Pinocembrin protects rat brain against oxidation and apoptosis induced by ischemia-reperfusion bothin vivoandin vitro[J].Brain Res,2008,1216:104 -15.
[11]Gao M,Zhang W C,Liu Q S,et al.Pinocembrin prevents glutamate-induced apoptosis in SH-SY5Y neuronal cells via decrease of bax/bcl-2 ratio[J].Eur J Pharmacol,2008,591(1-3):73 -9.
[12]Gao M,Liu R,Zhu S Y,Du G H.Acute neurovascular unit protective action of pinocembrin against permanent cerebral ischemia in rats[J].J Asian Nat Prod Res,2008,10(6):551 -8.
[13]Schacher S,Kandel E R,Montarolo P.cAMP and arachidonic acid simulate long-term structural and functional changes produced by neurotransmitters in Aplysia sensory neurons[J].Neuron,1993,10(6):1079-88.
[14]Glanzman D L,Kandel E R,Schacher S.Target-dependent structural changes accompanying long-term synaptic facilitation in Aplysia neurons[J].Science,1990,249(4970):799 - 802.
[15]Conrad P,Wu F,Schacher S.Changes in functional glutamate receptors on a postsynaptic neuron accompany formation and maturation of an identified synapse[J].J Neurobiol,1999,39(2):237-48.
[16]Hardingham G E,Bading H.Synaptic versus extrasynaptic NMDA receptor signalling:implications for neurodegenerative disorders[J].Nat Rev Neurosci,2010,11(10):682 - 96.
[17]Siegelbaum S A,Belardetti F,Camardo J S,Shuster M J.Modulation of the serotonin-sensitive potassium channel in aplysia sensory neuron[J].J Exp Biol,1986,124:287 - 306.
[18]Lodish H,Berk A,Zipursky S L,et al.Neurotransmitter receptors[M/OL].Molecular cell biology.4th edition.New York:W.H.Freeman,2000:Section 21.5.http://www.ncbi.nlm.nih.gov/books/NBK21586/.
