降解黃曲霉毒素枯草芽孢桿菌的解毒性、抗菌性及抗逆性研究
2011-06-08 03:15:44雷元培趙麗紅馬秋剛鄭文革高欣計成
飼料工業 2011年24期
雷元培 趙麗紅 馬秋剛 鄭文革 高欣 計成
黃曲霉毒素(AFT)主要是由黃曲霉(Aspergillusflavus)和寄生曲霉(Aspergillus parasiticus)產生的具有強烈毒性的次級代謝產物[1]。現已發現18種結構衍生物,其中黃曲霉毒素B1毒性最大,其具有極強的致癌、致突變、致畸和免疫抑制性[2-3]。黃曲霉毒素廣泛存在于玉米、花生粕等飼料原料中,嚴重威脅動物的生產性能,每年給飼料工業和畜牧業造成巨大的經濟損失[4-5]。自1960年被發現以來,AFT已經受到廣泛的重視,人們紛紛研究和采用各種方法來去除糧食和飼料中的AFT,其中包括各種物理、化學和微生物脫毒方法等[6-7]。然而,傳統的物理和化學去除霉菌毒素的方法存在效果不穩定、營養成分損失大、影響飼料適口性且難以規模化生產等缺點而不能被廣泛應用到實際生產中。微生物及生物酶解毒因具有解毒效率高、特異性強、對飼料和環境沒有污染等特點和優勢而備受研究者的重視[8-9]。
枯草芽孢桿菌(Bacillus subtilis)是一種嗜溫性好氧產芽孢革蘭氏陽性細菌,對人畜無毒、無害,不污染環境,具有廣譜抗菌活性和極強的抗逆能力,能分泌多種抗生素和酶。枯草芽孢桿菌的商品制劑是國際公認的可直接飼喂動物的、安全級(GRAS)微生物制品,對動物飼料酶制劑的開發有很好的應用前景。鑒于芽孢桿菌諸多功能特性,我們嘗試了從不同種類芽孢桿菌中篩選對黃曲霉毒素有降解活性的菌株,從35株實驗室分離并保藏的芽孢桿菌中得到一株對黃曲霉毒素有高效降解活性的菌,其發酵液對AFB1、AFG1和AFM1的降解率分別可達到81%、78%和63%。經16S rDNA鑒定為枯草芽孢桿菌(Bacillus subtilis),根據其功能特性命名為霉立解060菌。本試驗對篩選得到的能夠高效降解黃曲霉毒素B1、G1和M1的霉立解060菌的解毒活性、抗菌活性及抗逆性進行研究,為將該株具有多功能特性的枯草芽孢桿菌應用到動物實際生產中提供理論支持。
1 材料與方法
1.1 試驗菌株
霉立解060菌,經鑒定為枯草芽孢桿菌(Bacillus Subtilis),本實驗室從動物腸道分離得到并保藏,該株菌已經保藏在中國普通微生物菌種保藏管理中心(CGMCC),保藏號為CGMCC NO.3440。同時,該株菌已申請國家發明專利并獲授權(專利號:ZL 200910242938.3)。
沙門氏菌(Salmonella Typhimurium)、大腸桿菌(Escherichia Coli)、金黃色葡萄球菌(Staphylococcus Aureus),本實驗室保藏,作為試驗用指示菌。
枯草菌619(對黃曲霉毒素沒有降解作用的另一株枯草芽孢桿菌),本實驗室保藏,用于抑菌性和抗逆性對比試驗。
1.2 試驗試劑與設備
黃曲霉毒素 B1、G1、M1購于 Sigma 公司;LB 培養基(1000 ml):胰蛋白胨 10 g、酵母浸粉 5 g、NaCl 10 g,pH 值 7.0~7.2,蛋白酶 K。
1.3 試驗方法
1.3.1 霉立解060菌降解黃曲霉毒素活性組分的確定
采用50 ml/250 ml裝液量的LB培養基對霉立解060菌進行接種,在37℃、200 r/min條件下搖瓶培養24 h制得霉立解060菌發酵液。取5 ml發酵液,4℃離心20 min(8000 r/min),分離上清液與菌體,上清液于4℃備用;離心后的菌體用蒸餾水洗滌,再離心,取菌體加入5 ml蒸餾水制得菌懸液備用;以同樣的方法再制備一份菌體,對菌體進行破碎,然后溶于5 ml蒸餾水制成胞內液,備用。
對制得的等量的發酵液、上清液、菌懸液及胞內液對黃曲霉毒素B1、G1和M1的降解活性進行測定。取800 μl霉立解060菌發酵液,加入200 μl黃曲霉毒素B1(500 μg/kg),在37℃黑暗的培養箱中反應72 h;以等量的LB培養基加黃曲霉毒素B1為對照組;以同樣的方法向霉立解060菌發酵液中分別加入AFG1和AFM1進行解毒反應試驗,用HPLC及柱后光化學衍生的方法分別對AFB1、AFG1和AFM1的降解率進行測定。
1.3.2 霉立解060菌發酵液活性組分性質的研究
對霉立解060菌發酵上清液分別進行熱處理(100℃加熱10 min)和蛋白酶K處理(0.01 g/ml蛋白酶K與發酵上清液反應2 h)。按上述方法進行解毒反應試驗,研究解毒活性物質的性質。
1.3.3 黃曲霉毒素含量測定
黃曲霉毒素含量測定可分為3個步驟:萃取、凈化、檢測。首先使用甲醇:水(6:4)溶液對其進行萃取,然后使用免疫親和柱對樣品殘留毒素進行凈化提取(方法參照免疫親和柱使用說明書);最后用HPLC(柱后光化學衍生)對凈化提取得到的樣品進行檢測。
首先,應當制定促進科技發展方面的立法。比如,我國制定《科技促進法》、《科技人才促進法》等促進科技發展的法律,為科技發展提供制度環境。其次,應當完善已有的科技相關法律。比如,應當從促進科技發展的角度出發,完善《著作權法》、《專利法》、《商標法》、《網絡安全法》等,為科技創新保駕護航,同時維護科研人員的智慧成果,激發其創新的積極性。最后,制定和完善規范科技運行的法律。比如,我國可以制定《科技倫理法》,對科技工作者和科學研究活動提出倫理性要求,禁止他們從事有違社會倫理道德的科研活動。另外,還可以制定法律禁止科學研究用于違法的行為。
HPLC檢測條件為流動相甲醇:水=1:1;流速 1 ml/min;色譜柱 C18150 mm×4.6 mm,0.5 μm;激發波長360 nm,檢測波長440 nm;進樣量 20 μl。
AFB1降解率(%)=[(對照組AFB1含量-處理組AFB1含量)/對照組 AFB1含量]×100。
AFG1和AFM1與AFB1降解率的檢測方法相同。
1.4 霉立解060菌的抗菌性與抗逆性試驗
1.4.1 抗菌性試驗
指示菌懸液制備時,用接種針從斜面上挑取少量的指示菌,分別接種到營養肉湯培養基中,37℃培養24 h,調節菌懸液的濃度為109CFU/ml左右。
采用平板擴散的方法(Lyver等,1998;林東等,2001)[10-11],將霉立解060菌或枯草菌619用接種棒取少許分別點種到倒好的營養瓊脂平板中央,37℃培養24 h,然后把平板倒扣于盛一薄層氯仿的培養皿蓋上并熏蒸約30 min,以便殺死活細胞并能讓菌落固定于平板的表面,然后移去培養皿蓋并讓平板里的殘余氯仿徹底揮發20 min。在平板表面鋪一層剛培養好的指示菌培養液,均勻布滿后傾倒出剩余的菌液,晾干平板,置平板于37℃培養24 h,觀察點種的芽孢桿菌周圍抗菌圈的大小和清晰程度來判斷受試菌對指示菌的抗菌能力。
1.4.2 抗逆性試驗
芽孢懸液的制備方法:將霉立解060菌或枯草菌619培養液置于80℃水浴保溫15 min,離心收集芽孢,菌體用磷酸緩沖液洗滌兩次,最后復溶在緩沖液中即為芽孢懸液。
加熱處理試驗:分別取保存好的霉立解060菌或枯草菌619菌液5 ml注入到離心管中,采用分級稀釋,平板涂布。再將裝有剩余菌液的離心管置于80℃水浴鍋中加熱15 min,取加熱后的菌液進行逐級稀釋,平板涂布。最后將加熱前和加熱后的平板均在37℃條件下培養24 h,計算兩株芽孢桿菌加熱前后的活菌數。
模擬胃液的耐受性試驗:模擬胃液的制備借鑒Huang等(2004)[12]的方法。取0.5 ml的菌液加入4.5 ml的模擬胃液中,并迅速在振蕩器上充分混和,然后置于37℃培養箱靜置培養。分別在0、2和4 h的時候取出培養液并立即計數殘存芽孢的活菌數。
模擬膽鹽的耐受試驗:模擬膽鹽的制備借鑒Huang等(2004)[12]的方法。取0.5 ml的菌液加入4.5 ml的模擬膽鹽中,并迅速在振蕩器上充分混和,然后置于37℃培養箱靜置培養。分別在0、24 h的時候取出培養液并立即計數殘存芽孢的活菌數。
芽孢總數的測定:樣品用無菌生理鹽水10倍梯度稀釋,選擇合適的稀釋梯度用于計數。采用營養瓊脂培養基,取0.5 ml稀釋液涂布在平板上,在37℃培養24 h。
2 結果與分析
2.1 霉立解 060菌發酵上清液對 AFB1、AFG1和AFM1有較高的降解活性(見圖1)

圖1 霉立解060菌發酵液不同組分對黃曲霉毒素B1、G1和M1的降解率
由圖1可知,霉立解060菌發酵上清液對AFB1、AFG1和AFM1的降解活性最高,分別為73%、71%和59%;而菌懸液和胞內液對黃曲霉毒素B1、G1和M1的降解率不高,僅在10%左右。因此,初步認為霉立解060菌對黃曲霉毒素降解的活性物質是一種胞外分泌物,主要存在于其發酵后的上清液中。
2.2 熱處理和蛋白酶K處理后霉立解060菌上清液對 AFB1、AFG1和 AFM1降解活性(見圖 2)

圖2 霉立解060菌發酵后的上清液熱處理和蛋白酶K處理后對AFB1、AFG1和AFM1的降解率
由圖2可知,霉立解060菌發酵后的上清液經熱處理和蛋白酶K處理后其對黃曲霉毒素的降解率顯著降低,發酵后的上清液經熱處理后對AFB1、AFG1和AFM1的降解率分別降低為20%、15%和17%,經蛋白酶K處理后分別降低為34%、28%和19%。因此,可初步判斷霉立解060菌對黃曲霉毒素的降解活性物質是一種細胞所產胞外酶。
2.3 霉立解060菌抑菌性測定(見表1)

表1 霉立解060菌對大腸桿菌、雞白痢沙門氏菌、金黃色葡萄球菌的抑菌活性
由表1可知,霉立解060菌對大腸桿菌有顯著的抑制活性,其抑菌圈直徑為0.98 cm,而枯草菌619對大腸桿菌沒有抑制作用,抑菌圈直徑為0;盡管枯草菌619對雞白痢沙門氏菌和金黃色葡萄球菌抑菌圈直徑比霉立解060菌的大,但由于其沒有抑制大腸桿菌的活性,因此,霉立解060菌是一株對多種有害菌有抑制作用、抑菌性強的有益微生物。
2.4 霉立解060菌抗逆性測定
模擬胃液對霉立解060菌活性的影響見表2。由表2可知,在模擬胃液條件下,霉立解060菌活菌數比枯草菌619活菌數高;與0 h活菌數相比,霉立解060菌經2 h和4 h模擬胃液處理后活菌數沒有顯著變化,活菌數仍達到98.83%和97.95%,說明霉立解060菌對胃酸具有很強的耐受性。

表2 模擬胃液對霉立解060菌活性的影響
模擬膽鹽對霉立解060菌耐受性的影響見表3。由表3可知,在模擬膽鹽的條件下,霉立解060菌0 h和24 h的活菌數沒有顯著變化(P>0.05),模擬膽鹽處理24 h后其活菌數仍然能達到96.03%;而枯草菌619經膽鹽處理24 h后活菌數為0,對膽鹽沒有耐受能力,進一步說明霉立解060菌是一株對膽鹽具有很好耐受性的菌。

表3 霉立解060菌對膽鹽耐受性試驗結果
霉立解060菌對高溫耐受性試驗的研究結果見表4。由表4可知,霉立解060菌在80℃加熱15 min后活菌數沒有變化(P>0.05),其活菌數仍能達到98.69%;枯草菌619在經過80℃加熱后,活菌數為0,結果表明,霉立解060菌對80℃高溫具有很好的耐受性,適合作為動物用飼料添加劑。
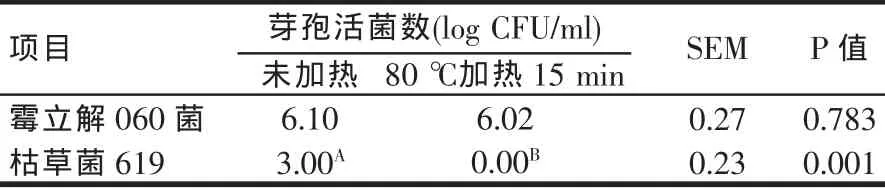
表4 霉立解060菌對高溫耐受性試驗結果
3 討論
霉立解060菌是課題組從動物腸道分離得到的,進一步研究表明,該株枯草芽孢桿菌對黃曲霉毒素降解的活性組分存在于發酵上清液中,菌懸液和胞內液對黃曲霉毒素的降解率很低。發酵上清液經熱處理和蛋白酶K處理后對黃曲霉毒素的降解率顯著降低,初步認為霉立解060菌對黃曲霉毒素的解毒活性物質是一種細胞所產胞外酶。眾所周知,枯草芽孢桿菌酶是工業酶市場的主體,其生產的蛋白酶、淀粉酶是工業酶中應用最為廣泛的酶,僅這兩種酶就占了整個工業酶市場的50%[13]。目前報道的能夠降解黃曲霉毒素的細菌不多,有橙色黃桿菌(Flavobacterium aurantiacum)[14]、分支桿菌(Mycobacterium fluoranthenivorans)[15]、紅串紅球菌(Rhodococcus erythropolis)[16]及橙紅色粘球菌(Myxococcus fulvus)[17]等。這些菌如果要應用到動物生產中還需要對其做進一步的安全性評價,而枯草芽孢桿菌是農業部允許直接添加到動物飼料中的有益微生物,因此芽孢桿菌霉立解060菌及其所產的毒素降解酶具有非常大的應用價值和廣闊的應用前景。
為進一步研究霉立解060菌的抑菌特性和抗逆活性,將該株菌應用到動物實際生產中,盡可能地發揮該菌的功能特性,我們對霉立解060菌和另外一株對黃曲霉毒素沒有降解作用的枯草芽孢桿菌進行對比試驗研究。動物腸道環境復雜,腸道菌群多樣性變化,除附殖有益微生物(乳酸菌、芽孢桿菌、腸球菌)外,還包含有害微生物,如大腸桿菌、雞白痢沙門氏菌、金黃色葡萄球菌等,進入動物腸道的有益微生物若能較好的發揮其功能,必須要抵抗各種不利因素,包括抵抗有害菌的抑制作用。通過以這三株菌為指示菌,以另外一株對黃曲霉毒素沒有解毒活性的枯草菌619做對比,研究了霉立解060菌的抑菌活性,結果表明,霉立解060菌對大腸桿菌、雞白痢沙門氏菌和金黃色葡萄球菌有較好的抑菌活性,可以抵抗動物腸道有害菌的不利影響而發揮正常的生理功能。試驗結果還表明,并不是所有的枯草芽孢桿菌都對多種有害菌同時具有抑菌活性。
目前,有益微生物均以口服方式飼喂動物,因此,它們必須首先具備耐受胃的低pH值的能力才可能進入動物腸道存活、繁殖,發揮其作用。除應具備耐胃低pH值能力外,還需具備耐受小腸中膽鹽等形成的高滲透壓環境的能力。枯草芽孢桿菌等益生菌株在菌液濃縮或制作膨化顆粒料時,都要進行加熱處理,對高溫的耐受性也成為選擇有益微生物的必備條件[18]。因此,試驗對霉立解060菌株進行了模擬胃液、膽鹽及熱耐受試驗。結果表明,與另外一株對黃曲霉毒素沒有解毒活性的枯草芽孢桿菌相比,霉立解060菌除具有降解黃曲霉毒素的特性外,還能顯著抑制動物腸道有害菌的繁殖,同時抵抗胃酸、膽鹽、高溫等不利環境而正常生長。因此,霉立解060菌具備良好的開發前景和應用價值。
4 結論
課題組從動物腸道分離篩選到的枯草芽孢桿菌(霉立解060菌)對黃曲霉毒素B1、G1和M1具有較高的降解活性;初步試驗證明,解毒活性物質是該菌分泌的一種胞外蛋白酶;同時,霉立解060菌具有很好的抑菌活性和抗逆性,非常適合用于動物飼料微生態制劑的開發和應用。
[1]Cotty P J,Bhatnagar D.Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic pathway enzymes[J].Applied and Environmental Microbiology,1994,60(7):2248-2251.
[2]Alberts J F,Engelbrecht Y,Steyn P S,et al.Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures[J].International Journal of Food Microbiology,2006,109(1/2):121-126.
[3]Pitt J I,Basilico J C,Abarca M L,et al.Mycotoxins and toxigenic fungi[J].Medical Mycology,2000,38(SUPPL.1):41-46.
[4]Massey T E,Stewart R K,Daniels J M,et al.Biochemical and molecular aspects of mammalian susceptibility to aflatoxin B1 carcinogenicity[J].Proceedings of the Society for Experimental Biology and Medicine,1995,208(3):213-227.
[5]計成.霉菌毒素與飼料食品安全[M].北京:化學工業出版社,2007.
[6]王寧,馬秋剛,張建云,等.黃曲霉毒素的傳統去毒方法和生物降解研究進展[J].飼料與畜牧,2008(7):18-20.
[7]Desheng Q,Fan L,Yanhu Y,et al.Adsorption of aflatoxin B1 on montmorillonite[J].Poultry Science,2005,84(6):959-961.
[8]Kabak B,Dobson A D W,Var I.Strategies to prevent mycotoxin contamination of food and animal feed:A review[J].Critical Reviews in Food Science and Nutrition,2006,46(8):593-619.
[9]Schatzmayr G,Zehner F,T Ubel M,et al.Microbiologicals for deactivating mycotoxins[J].Molecular Nutrition and Food Research,2006,50(6):543-551.
[10]Lyver A,Smith J P,Austin J,et al.Competitive inhibition of Clostridium botulinum type E by Bacillus species in a valueadded seafood product packaged under a modified atmosphere[J].Food Research International,1998,31(4):311-319.
[11]林東,徐慶,劉憶舟,等.枯草芽孢桿菌SO113分泌蛋白的抑菌作用及抗菌蛋白的分離純化 [J].農業生物技術學報,2001,9(1):77-80.
[12]Huang Y,Adams M C.In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propionibacteria[J].International Journal of Food Microbiology,2004,91(3):253-260.
[13]Schallmey M,Singh A,Ward O P.Developments in the use of Bacillus species for industrial production[J].Canadian Journal of Microbiology,2004,50(1):1-17.
[14]Ciegler A,Lillehoj E B,Peterson R E,et al.Microbial detoxification of aflatoxin[J].Applied Microbiology,1966,14:934-939.
[15]Hormisch D,Brost I,Kohring G W,et al.Mycobacterium fluoranthenivorans sp.nov.,a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant[J].Systematic and Applied Microbiology,2004,27(6):653-660.
[16]Teniola O D,Addo P A,Brost I M,et al.Degradation of aflatoxin B1 by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp.nov.DSM44556T[J].International Journal of Food Microbiology,2005,105(2):111-117.
[17]Zhao L H,Guan S,Gao X,et al.Preparation,purification and characteristics of an aflatoxin degradation enzyme from Myxococcus fulvus ANSM068 [J].Journal of Applied Microbiology,2011,110(1):147-155.
[18]劉虹,姚文,于卓騰,等.一組雞源乳酸菌產乳酸及其耐受特性研究[J].微生物學通報,2006(5):1-5.
