烏蘇里江秋大麻哈魚生殖洄游群體結(jié)構(gòu)特征與分析
2011-06-08 07:53:56王繼隆唐富江潘志強(qiáng)
湖南農(nóng)業(yè)科學(xué) 2011年21期
王繼隆,唐富江 ,朱 翥 ,潘志強(qiáng) ,劉 偉
(1.中國水產(chǎn)科學(xué)研究院黑龍江水產(chǎn)研究所,黑龍江 哈爾濱 150070;2.農(nóng)業(yè)部黑龍江流域漁業(yè)資源環(huán)境科學(xué)觀測實(shí)驗站,黑龍江 哈爾濱 150070;3.黑龍江省撫遠(yuǎn)縣鮭魚放流站,黑龍江 撫遠(yuǎn) 156500)
大麻哈魚(Oncorhynchus keta Walbaum)是典型的溯河洄游魚類,幼魚春季降海、成魚秋季溯河洄游至出生地產(chǎn)卵直至生命終結(jié),終生只繁殖一次。其廣泛分布于白令海、北太平洋、鄂霍次克海、日本海及沿岸河流中[1],其中加拿大、美國、日本、俄羅斯數(shù)量分布較多。中國境內(nèi)的大麻哈魚可分為4個地方種群,即黑龍江、烏蘇里江、綏芬河、圖們江[2]。在烏蘇里江大麻哈魚幼魚春節(jié)隨水流降海,最終在黑龍江河口進(jìn)入鄂霍次克海,而后在西太平洋、白令海等進(jìn)入海洋生活階段,直至性成熟后上溯至出生地產(chǎn)卵繁殖。
國外學(xué)者對大麻哈魚洄游群體生物學(xué)特性進(jìn)行了較深入的研究[3-5],上世紀(jì)90年代學(xué)者研究發(fā)現(xiàn)太平洋大麻哈魚個體小型化及成熟年齡增大的現(xiàn)象,認(rèn)為影響因素包含環(huán)境因子、捕撈選擇性和種群壓力等[6-10]。近年來日本學(xué)者研究發(fā)現(xiàn)大麻哈魚小型化現(xiàn)象主要受到種間競爭的影響[11-12]。中國對大麻哈魚資源調(diào)查也開展了一些研究,成熟個體小型化等現(xiàn)象也逐漸被揭示[13-15]。但對大麻哈魚資源調(diào)查尚缺乏持續(xù)性和深入性,尤其是近幾十年來大麻哈魚資源增殖放流工作的不斷開展,同時伴隨著捕撈強(qiáng)度的居高不下,大麻哈魚資源及種群結(jié)構(gòu)變動趨勢更加難以預(yù)料。筆者對2010年烏蘇里江大麻哈魚回歸群體捕撈情況進(jìn)行了持續(xù)監(jiān)測,并對其回歸群體數(shù)量結(jié)構(gòu)進(jìn)行分析,旨在了解大麻哈魚洄游群體數(shù)量結(jié)構(gòu)及生物學(xué)特征,為研究大麻哈魚資源提供參考。
1 材料與方法
1.1 調(diào)查時間及區(qū)域
該研究在大麻哈魚禁漁期(2010年10月1日到2010年10月20日)跟蹤調(diào)查6艘增殖放流親魚采捕船只作業(yè)情況。禁漁期增殖放流捕撈船只數(shù)量固定,且作業(yè)地點(diǎn)也統(tǒng)一集中在通江口江段(其他江段禁捕)。
烏蘇里江是一條中俄界河,發(fā)源于俄羅斯遠(yuǎn)東地區(qū)的錫霍特山脈,自南向北于黑瞎子島地區(qū)注入黑龍江,最終注入鄂霍次克海。洄游期大麻哈魚從黑龍江河口上溯至黑、烏交界處,然后分別游向黑龍江和烏蘇里江。研究調(diào)查地點(diǎn)位于黑龍江省撫遠(yuǎn)縣通江口江段(48.3°N,134.7°E)(圖 1),也是中國境內(nèi)大麻哈魚洄游至烏蘇里江的起點(diǎn)。
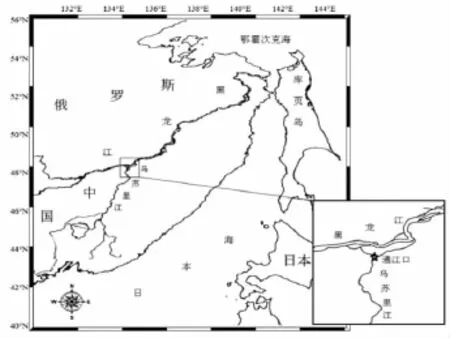
圖1 調(diào)查站點(diǎn)
1.2 數(shù)據(jù)收集
大麻哈魚捕撈網(wǎng)具為3層流刺網(wǎng),網(wǎng)目大小為12.67 cm,網(wǎng)高3.9m,長約500m,作業(yè)時網(wǎng)具隨水漂流,同時網(wǎng)具下綱觸底。由于大麻哈魚捕撈船大小、主機(jī)功率及網(wǎng)具較為統(tǒng)一,所以單位捕撈努力量漁獲量(CPUE)統(tǒng)一為kg/(船·日),調(diào)查期間逐日統(tǒng)計了6艘捕撈船只捕獲雌、雄大麻哈魚質(zhì)量。在調(diào)查期間每天隨機(jī)收集一部分大麻哈魚樣本,共采集樣本505尾,其中雄魚237尾,雌魚268尾,測量體長、體質(zhì)量,并采集背鰭末端至臀鰭之間且側(cè)線上方6~10枚鱗片供年齡鑒定。
1.3 處理方法
對6條漁船的雌、雄大麻哈魚產(chǎn)量進(jìn)行平均值處理,并根據(jù)雌雄個體平均質(zhì)量換算為雌雄個體數(shù)量。然后對收集的大麻哈魚樣本進(jìn)行測量,分別統(tǒng)計雌雄個體體長、體質(zhì)量、年齡等生物學(xué)指標(biāo),并且通過方差分析判斷不同洄游時期的大麻哈魚的生物學(xué)指標(biāo)的差異性。
2 結(jié)果與分析
2.1 回歸群體數(shù)量特征
由于10月1日和10月20日捕撈船只的作業(yè)時間和其他日期作業(yè)時間不一致,故未將其統(tǒng)計在內(nèi)。回歸群體的數(shù)量特征反映了大麻哈魚的群體結(jié)構(gòu)特點(diǎn),具體特征如下。
從圖2可以看到禁漁期開始時回歸群體的數(shù)量最多,10月9日處于低潮期,10月2日到9日基本成遞減趨勢,10日到15日成遞增趨勢,而后基本呈波動下降趨勢,直至漁期結(jié)束。該調(diào)查發(fā)現(xiàn)9月25日大麻哈魚開始形成漁汛,隨后回歸數(shù)量增減增多。由于缺乏9月25日到10月1日的數(shù)據(jù),未能給出整個漁期的數(shù)量結(jié)構(gòu)圖,但從中也可看出大麻哈魚回歸數(shù)量出現(xiàn)兩個波峰,即10月初出現(xiàn)一次大高峰,10月15日出現(xiàn)一次小高峰。禁漁期過后大麻哈魚數(shù)量很少,已不能形成漁汛。因此,根據(jù)大麻哈魚洄游數(shù)量結(jié)構(gòu)將洄游期分為兩個階段,即10月10日前為第一階段,10月10日后為第二階段。
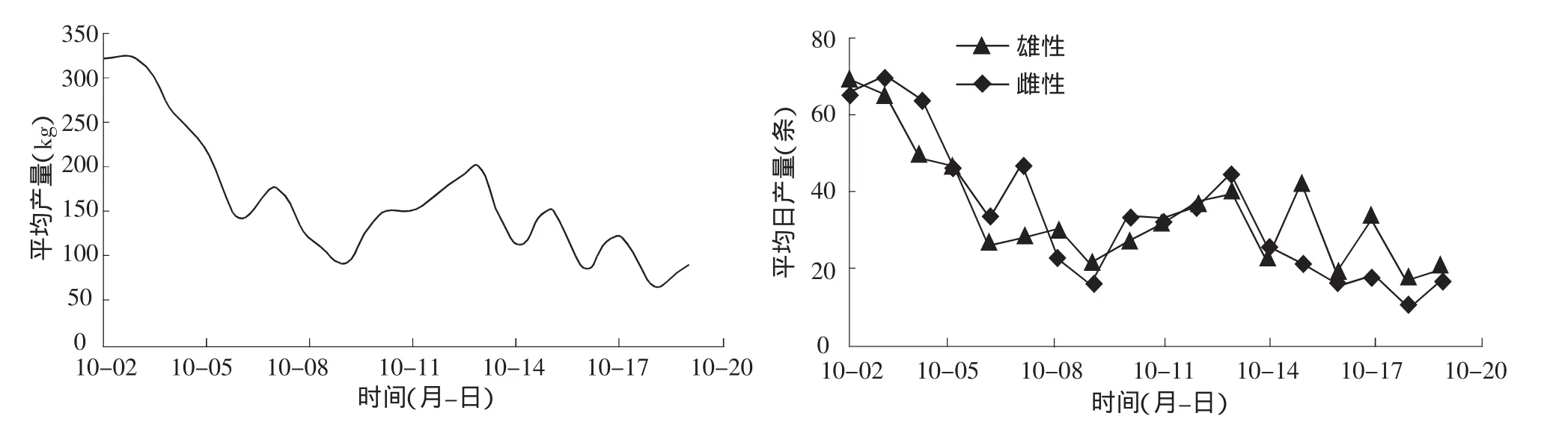
圖2 大麻哈魚日均產(chǎn)量及雌雄個體數(shù)量分布
在整個調(diào)查期內(nèi)大麻哈魚雌、雄個體數(shù)量比例為1.01∶1。從圖2也可以看出,雌、雄大麻哈魚的數(shù)量以及和總捕撈量變化趨勢基本一致,第一階段和第二階段大麻哈魚雌、雄個體間數(shù)量比例變動是隨機(jī)的。
2.2 不同洄游期階段群體間的差異性分析
對洄游期兩個階段的大麻哈魚的各項生物學(xué)指標(biāo)(性比、叉長、體質(zhì)量和年齡)進(jìn)行了方差分析(表1),方差分析結(jié)果p值均大于0.05,表明各指標(biāo)差異性均不顯著。
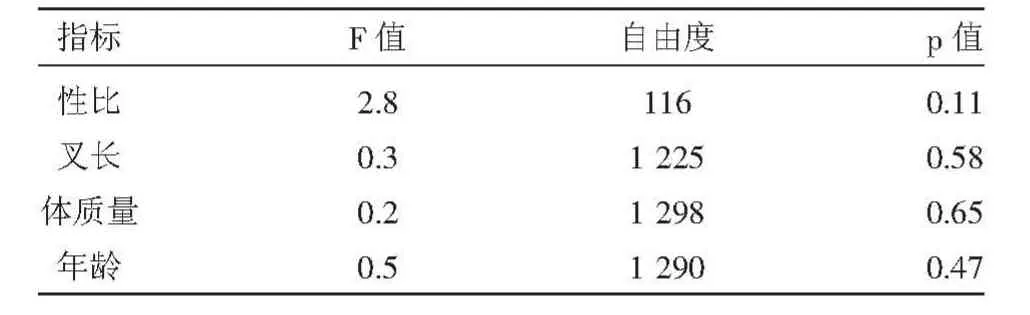
表1 兩洄游階段的大麻哈魚各項生物學(xué)指標(biāo)的方差分析
2.3 大麻哈魚洄游群體的生物學(xué)特征
2.3.1 年齡組成 總體上說,大麻哈魚年齡范圍為2+~4+齡,其中 2+占 42.8%、3+占 53.4%、4+占 3.8%,可見3+為優(yōu)勢年齡組。方差分析表明雌雄群體間的年齡組成差異顯著(p<0.01)。雄性個體中2+占53.4%、3+占 42.7%、4+占 3.8%,2+為優(yōu)勢年齡組;雌性個體中2+占 34.2%、3+占 62.1%、4+占 3.7%,3+為優(yōu)勢年齡組。可見雄性大麻哈魚成熟期比雌性要短。

表2 大麻哈魚的年齡組成 (%)
2.3.2 體長、體質(zhì)量 大麻哈魚雌、雄性個體的叉長、體長、體質(zhì)量情況如表3所示,雄性個體平均叉長、體長、體質(zhì)量分別為60.6 cm、63.5 cm、2 309.5 g;雌性個體平均叉長、體質(zhì)量分別為62.4 cm、65.5 cm、2 532.3 g。可見,雄性個體平均體長和體質(zhì)量都小于雌性個體。雄性個體叉長、體質(zhì)量主要集中在 58~64 cm、2 150~2 750 g;雌性個體叉長、體質(zhì)量主要集中在 59~65 cm、2 250~2 750 g。

表3 雌、雄大麻哈魚的體長、叉長、體質(zhì)量分布
3 討論
3.1 大麻哈魚生殖洄游群體數(shù)量特征
根據(jù)大麻哈魚回歸群體數(shù)量結(jié)構(gòu)特征,將洄游期分為10月10日前、后的兩個階段。而以往的研究將這兩個階段統(tǒng)一起來,沒有深入?yún)^(qū)分[13,15],這種分期現(xiàn)象更能詳細(xì)反映實(shí)際捕撈狀況。不同洄游階段的大麻哈魚的各項生物學(xué)指標(biāo)方差分析表明其差異性并不顯著,這也說明不同洄游期間的大麻哈魚只是數(shù)量結(jié)構(gòu)上的變化,生物學(xué)指標(biāo)沒有明顯差異。因此,這種數(shù)量的波動可能是由不同洄游群體間的分散效應(yīng)造成。此外,俄羅斯在黑龍江河口對大麻哈魚回歸群體也有一定的捕撈,其對洄游至中國烏蘇里江的大麻哈魚數(shù)量有直接影響。
3.2 洄游群體生物學(xué)特征
3.2.1 成熟年齡低齡化 20世紀(jì)60年代以來,洄游至中國大麻哈魚的性成熟平均年齡呈現(xiàn)是在波動中下降趨勢(圖3),2010年大麻哈魚性成熟年齡較近幾年呈下降趨勢[13],并且達(dá)到歷史最低值。90年代國外有研究發(fā)現(xiàn)大麻哈魚性成熟年齡有升高的趨勢[16-17]。大麻哈魚種群分布的不同也會造成不同的結(jié)果[18]。對成熟年齡變化的原因研究相對較少,一直沒有明確的結(jié)論。漁業(yè)捕撈的持續(xù)選擇性會造成大麻哈魚性成熟年齡的降低[18-23],這是因為高年齡魚個體大,被捕撈的風(fēng)險也較大,所以漁業(yè)捕撈的長期選擇性會使得性成熟年齡降低,但是成熟年齡大小的波動卻不能完全解釋。
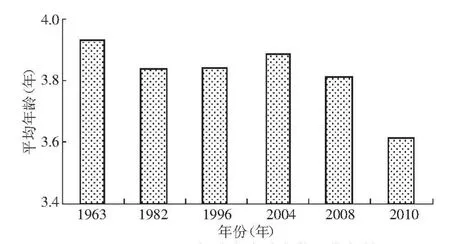
圖3 不同年份大麻哈魚的平均年齡
有學(xué)者通過模型模擬出大麻哈魚性成熟年齡和生長速度之間的負(fù)相關(guān)關(guān)系[24]。20世紀(jì)90年代國外學(xué)者分析了大麻哈魚性成熟年齡變大和體長減小的現(xiàn)象[10,16]時,指出這是減緩生長速度應(yīng)對環(huán)境變化的響應(yīng)[22]。根據(jù)這種理論,近年來烏蘇里江大麻哈魚成熟年齡的降低是由于生長速度變快造成的,同樣可以解釋圖3中成熟年齡的波動原因。然而在生態(tài)系統(tǒng)中影響大麻哈魚生長速度的因素非常復(fù)雜,有遺傳因素和棲息地環(huán)境等因素共同影響。3.2.2 群體結(jié)構(gòu)小型化 2010年烏蘇里江秋大麻哈魚生殖洄游群體平均體長、體質(zhì)量與歷史資料相比呈減小趨勢,這也順延了20世紀(jì)60年代以來的總體下降趨勢(圖4)。一方面是因為群體年齡結(jié)構(gòu)的降低導(dǎo)致群體結(jié)構(gòu)小型化;另一方面就是生活環(huán)境導(dǎo)致魚類個體小型化,由于缺乏歷史資料,只能對此簡單探討。
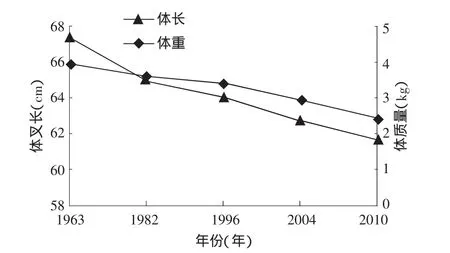
圖4 烏蘇里江大麻哈魚不同年份的平均叉長和體質(zhì)量
國內(nèi)外對太平洋大麻哈魚體長小型化現(xiàn)象也早有報道[16-18]。針對魚類小型化現(xiàn)象的原因,有研究發(fā)現(xiàn)和環(huán)境有相關(guān)關(guān)系,如溫度、鹽度等。但是成熟大麻哈魚體長的連續(xù)下降不能完全用環(huán)境變化來解釋,捕撈選擇性所引起種群遺傳結(jié)構(gòu)的變化而造成的成熟群體小型化也是不容忽視的[7,19-20],捕撈活動引起的高死亡率和個體大小的選擇性是引起魚類個體大小遺傳進(jìn)化的兩個因素[21-22]。隨著太平洋大麻哈增殖放流活動力度的不斷增強(qiáng),大麻哈魚群體數(shù)量的不斷增加,由此帶來種間競爭壓力的增大也是導(dǎo)致魚體小型化的因素之一。
據(jù)了解,中國在20世紀(jì)60年代曾在烏蘇里江對大麻哈魚進(jìn)行過增殖放流,之后基本上停止了這項活動,而俄羅斯對大麻哈魚放流也主要集中在黑龍江流域。因此,目前中國境內(nèi)烏蘇里江大麻哈魚基本上是天然種群。同時由于烏蘇里江大麻哈魚種群數(shù)量相對較少,在海洋中和其他大麻哈魚種群地理分布不同[,18],種間壓力相對較小。因此,種間競爭不應(yīng)是大麻哈魚小型化的主要原因。而中國大麻哈魚的捕撈強(qiáng)度一直保持在較高水平,捕撈死亡率大于可開發(fā)率從而造成漁業(yè)資源的衰退,同時捕撈網(wǎng)具對個體大的大麻哈魚選擇性較高,小個體魚類逃逸的可能性要大于大個體種類。因此,在中國捕撈選擇性應(yīng)該是造成大麻哈魚小型化的主要原因。
綜上所述,大麻哈魚成熟期體長、體質(zhì)量、年齡的變化是環(huán)境因子、生物間的相互作用、選擇性捕撈的綜合作用的結(jié)果。在生態(tài)系統(tǒng)中,魚類小型化、成熟年齡變化可以說是對周圍環(huán)境的一種響應(yīng),由于生態(tài)系統(tǒng)復(fù)雜性,很難確切說明這些生物學(xué)特征變化的具體原因。因此,這方的研究需漁業(yè)生態(tài)研究學(xué)者的共同努力。
[1] Shigehiko U,Tomonori A,Penelope A,et al.Origin and Distribution of Chum Salmon in the Bering Sea during the Early Fall of 2002:Estimates by Allozyme Analysis[R].National Salmon Resources Center,Toyohira-ku,Sapporo 062-0922,Japan,2004.
[2] Chen JP,Sun D J,Dong C Z,et al.Genetic analysis of four wild chum salmon Oncorhynchus keta populations in China based on microsatellitemarkers[J].Environmental Biology of Fishes.2005,(73):181-188.
[3] Hideji T,Yasuaki T,Yasuhiko N.Behavioural thermoregulation of chum salmon during homingmigration in coastalwaters[J].The Journal of Experimental Biology.2000,(203):1825-1833.
[4] Manu E.Observations of spawning behaviour in Salmoninae:Salmo,Oncorhynchus and Salvelinus[J].Fish Biology and Fisheries.2005,(15):1-21.
[5] Manu E.Two examplesof fixed behavioural patterns in salmonines female false spawning and male digging[J].Japan Ethological Society and Springer-Verlag.2007,(25):63-70.
[6] Brian SB,David W W,John H H.A review of size trends among North PacificA review of size trends among North Pacific salmon(Oncorhynchus spp.)[J].Canadian Journal of Fisheries and Aquatic Sciences.1996,(53):455-465.
[7] Richard L.Fishing selection and phenotypic evolution[J].Journal of Marine Science.2000,(57):659-668.
[8] Shoko H M,Kentaro M,Hiroyuki S.Growth of chum salmon(Oncorhynchus keta)correlated with sea-surface salinity in the North Pacific[J].Journal of Marine Science.2001,(58):1335-1339.
[9] Ishida Y,Welch DW,Ogura M.Potential influence of North Pacific sea-surface temperatures on increased production of chum salmon(Oncorhynchus keta)from Japan[J].Climate change and northern fish populations.1995,(121):271-275.
[10] Ishida Y,Ito S,Kaeriyama M,et al.Recent changes in age and size of chum salmon (Oncorhynchus keta)in the North Pacific Ocean and possible causes[J].Fisheries and Aquatic Sciences.1993,(50):290-295.
[11] Hyunju S,Hideaki K,Masahide K.Spatiotemporal change in growth of two populations of Asian chum salmon in relation to intraspecific interaction[J].Fish Sci.2009,(75):957-966.
[12] Hyunju S,Hideaki K,Masahide K.Long-term climate-related changes in somatic growth and population dynamics of Hokkaido chum salmon[J].Environ Biol Fish.2011,(90):131-142.
[13] 董崇智,趙春剛,王 金,等.烏蘇里江大麻哈魚的溯河生殖群體結(jié)構(gòu)[J].中國水產(chǎn)科學(xué),1999,6(3):5-9.
[14] 趙春剛,潘偉志,陳 軍,等.2004年烏蘇里江大麻哈魚生殖群體結(jié)構(gòu)研究[J].水產(chǎn)學(xué)雜志,2005,18(2):59-64.
[15] 唐富江,劉 偉,徐鳳龍,等.黑龍江水系(中國)秋大麻哈魚回歸群體變動分析[J].上海海洋大學(xué)學(xué)報,2010,19(1):74-79.
[16] Kaev A.Dynamics of some biological indexes of Oncorhynchus keta in connection with the formation of its numbers[J].Journal of ichthyology.1999,39(88):642-651.
[17] John H H,Margaret SH.Size decline and older age atmaturity of two chum salmon(Oncorhynchus keta)stocks in western North America[J].Fish Aquature.1995,(121):245-260.
[18] Shogo M,Shunpei S,Moongeun Y,et al.Nonrandom distribution of chum salmon stocks in the Bering Sea and the North Pacific Ocean estimated using mitochondrial DNA microarray[J].Fish Science.2009,(75):359-367.
[19] Ricker W E.Changes in the average size and average age of Pacific salmon[J].Journal of Fisheries and Aquatic Sciences.1981(38):1636-1656.
[20] David O C,Stephan BM.Sustaining Fisheries Yields Over Evolutionary Time Scales[J].Science.2002,297(5578):94-96.
[21] Terry D B,Clyde B M.A genetic analysis of body size in pink salmon(Oncorhynchus gorbuscha)[J].Genome.1988,(30):31-35.
[22] Kentaro M,Masa-aki F.Why age and size at maturity have changed in pacific salmon[J].Marine Ecology Progress Series.2007,(335):289-294.
[23] Jorgensen T.Long-term changes in age at sexual maturity of Northeast Arctic cod(Gadusmorhua L.)[J].Conseil International pour l'Exploration de la Mer.1990,(46):235-248.
[24] Morita K,Morita S,Fukuwaka M,et al.Rule of age and size at maturity of chum salmon(Oncorhynchus keta):implications of recent trends among Oncorhynchus spp.[J].Canadian Journal of Fisheries and Aquatic Sciences.2005,(62):2752-2759.
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
現(xiàn)代畜牧科技(2021年4期)2021-07-21 06:13:00
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
福建基礎(chǔ)教育研究(2019年1期)2019-09-10 07:22:44
中國生殖健康(2019年2期)2019-08-23 08:12:08
福建基礎(chǔ)教育研究(2019年1期)2019-05-28 08:39:49
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
數(shù)學(xué)物理學(xué)報(2017年2期)2017-06-05 09:12:30
汽車觀察(2016年3期)2016-02-28 13:16:26
醫(yī)學(xué)研究雜志(2015年11期)2015-06-10 06:44:03
