嚙齒動物種群數量與年降水量的灰色關聯度分析
2011-03-31 06:12:56包俊江武曉東付和平張福順李穎班曉瑞
草業科學 2011年4期
關鍵詞:研究
包俊江,武曉東,付和平,張福順,李穎,班曉瑞
(1.內蒙古農業大學生態環境學院,內蒙古 呼和浩特 010019;2.內蒙古工業大學能源與動力工程學院,內蒙古 呼和浩特 010062;3.內蒙古鄂爾多斯市氣象局,內蒙古 鄂爾多斯 017000;4.內蒙古自治區煙草質量監督檢測站,內蒙古 呼和浩特 010010)
嚙齒動物種群數量和氣候因子之間的關系一直是生態學家們關心的問題,尤其以北美、南美和澳大利亞地區由El Nio現象引起的降水量變化對嚙齒動物的影響研究較多[1-2],但所得結論并不一致。有研究認為嚙齒動物數量不受降水變化的影響[3],而有研究表明嚙齒動物的數量暴發與降水高峰是相關的[4]。這些研究往往僅對單一物種進行分析[5-7],缺少對多個物種的綜合研究。
阿拉善荒漠是內蒙古西部典型的溫性荒漠,嚙齒動物作為荒漠草地中重要的功能群,其數量大小和生態功能對生態系統的平衡和穩定具有重要意義。降水稀少而且極不均勻是荒漠生態系統的主要特點[8],對嚙齒動物種群數量大小存在直接或間接作用。本研究地區天然草地存在圍欄禁牧、適度輪牧和過度放牧3種利用方式,但劃區輪牧是該地區的主要利用方式,輪牧草地面積較大。因此,本研究對輪牧區中嚙齒動物和降水量的相關性進行分析。降水量與嚙齒動物種群數量之間的關系是非線性的[9],運用線性分析方法很難揭示事物的內在聯系。而灰色關聯度分析主要是通過對灰色系統中不同事物間的相關分析,根據因素之間發展趨勢的相似或相異程度,衡量因素間關聯程度的一種非線性方法[10]。本研究采用該方法計算群落中主要鼠種種群數量與年降水量的關聯度大小,分析各鼠種對降水量變化的反應,為揭示嚙齒動物群落格局變動規律提供依據。
1 研究區概況與研究方法
1.1 研究區自然概況研究區位于內蒙古阿拉善左旗南部的嘉爾格勒賽汗鎮,地理坐標為104°10′~105°30′E,37°24′~38°25′N,地處騰格里沙漠東緣。該地區的草地類型是典型的溫性荒漠,植被稀疏,結構單調,覆蓋度低,一般為1%~20%。植物種類貧乏,主要以旱生、超旱生和鹽生的灌木和小半灌木為主。建群植物以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygophyllaceae)為主,其次為薔薇科(Rosaceae)、檉柳科(Tamaricaceae)。地形起伏不平,丘陵、沙丘與平灘相間。氣候為典型的高原大陸性氣候,冬季嚴寒、干燥,夏季酷熱,晝夜溫差大,極端最低氣溫-36℃,最高氣溫42℃,年平均氣溫8.3℃,無霜期156 d。年降水量45~215 mm,且降水極不均勻,主要集中在7-8月。年蒸發量3 000~4 700 mm。土壤為棕漠土,淋溶作用微弱,土質松散、瘠薄,表土有機質含量1.0%~1.5%,含有較多的可溶性鹽。
1.2 研究方法2000年,研究區域開始實行劃區輪牧制度,2002年選擇500 hm2的天然草地作為試驗區,劃分為 4個輪牧小區,放牧 300只羊,各小區每月放牧一次。為了保證結論的準確性,研究期間,沒有更換試驗區,并且放牧制度始終保持一致。
2002-2008年每年的4、7、10月的上旬采用鋏日法對嚙齒動物種群數量進行調查。每次取樣時,在每個輪牧小區隨機選擇1個鋏捕樣方,樣方面積10 hm2。每個樣方布設5條鋏線,每鋏線100鋏,鋏距5 m,鋏線距50 m,共計500鋏日。以嚙齒動物的捕獲率代表種群的相對密度,用各年度4、7、10月的平均捕獲率表示該年度的捕獲率。2002-2008年試驗區的降水量數據來源于內蒙古阿拉善盟孿井灘氣象站。
1.3 分析方法采用灰色系統分析方法中的灰色關聯分析。其原理是依據空間理論為數學基礎,按照規范性、偶對稱性、整體性和接近性的原則,對信息部分確定和部分不確定的灰色系統的發展勢態進行定量描述和比較,以確定參考數列和若干比較數列之間的灰色關系。具體分析步驟如下。
將降水量與各鼠種的種群密度視為一個整體,即灰色關聯系統。并將降水量設定為參考數列X0,記作:X0={X0(1),X0(2),…,X0(n)},捕獲鼠種的種群密度分別設定為比較數列X1~Xi,記作:Xi={Xi(1),Xi(2),…,Xi(n)},(i=1,2,…,n),n為鼠種數。對各數列進行均值化無量綱處理,即將同一個數列的所有數據均除以該序列均值得到一個新數列,計算公式表示為:

求出參考數列X0與比較數列Xi各對應點的絕對差值,即 Δi(k),

關聯系數的計算:

關聯度(ri)的計算:

式中,ri為比較數列與參考數列的關聯度,是具體反映X0與Xi數列之間關聯性的度量[7]。
2 結果與分析
2.1 嚙齒動物群落的物種組成2002-2008年共布放 42 000鋏日,捕鼠2 861只(表1),分屬4科11種。它們分別為跳鼠科(Dipodidae):三趾跳鼠(Dipussagitta)、五趾跳鼠(Allactaga sibirica)、蒙古羽尾跳鼠(Stylodipus andrewsi)、三趾心顱跳鼠(Salpingotus kozlovi);倉鼠科(Cricetidae):小毛足鼠(Phodopus roborovskii)、黑線倉鼠(Cricetulusbarabensis)、短耳倉鼠(Allocricetuluseversmanni)、子午沙鼠(Meriones meridianus)、長爪沙鼠(M.unguiculatus);松鼠科(Sciuridae):阿拉善黃鼠(Spermophilus alaschanicus);鼠科(Muridae):小家鼠(Musmusculus)。子午沙鼠、三趾跳鼠、小毛足鼠、五趾跳鼠、阿拉善黃鼠和黑線倉鼠為群落中的主要鼠種(表1),各年中的捕獲數量較大,所以本研究僅對以上鼠種進行了種群數量和降水量的灰色關聯度分析。其中子午沙鼠和小毛足鼠為群落的優勢種,7年中的捕獲量明顯高于其他鼠種。

表1 2002-2008年嚙齒動物群落的物種組成 只
2.2 嚙齒動物種群數量與降水量的年動態2002-2008年群落中總的嚙齒動物數量與年降水量的變動規律如圖1所示,嚙齒動物種群數量和降水量的年動態波動均較大,變動趨勢基本一致。嚙齒動物的種群數量在2003年達到一個數量高峰,隨后降低,2006達到最低點,總捕獲率僅為1.18%,隨后逐漸上升,2008年達到另一個數量高峰,總捕獲率高達21.02%。降水量的第1個高峰也出現2003年,2005年降至最低,僅為63.3mm,隨后上升,2007年達到另一高峰,為288 mm。2002-2008年嚙齒動物的捕獲率與年降水量的變動趨勢是一致的,僅2008年有差異,2008年降水量較2007年降低,而嚙齒動物的捕獲率明顯高于2007年。但是經Spearman相關分析發現,2002-2008年總的嚙齒動物數量與年降水量呈顯著正相關(r=0.390 5,P=0.032 4)。

圖1 嚙齒動物捕獲率和降水量的年動態
2.3 嚙齒動物種群數量與年降水量的灰色關聯度分析
2.3.1數據無量綱化處理 將年降水量設定為參考數列X0,五趾跳鼠、三趾跳鼠、阿拉善黃鼠、小毛足鼠、子午沙鼠和黑線倉鼠的年捕獲率分別設定為比較數列X1~X6。由于嚙齒動物捕獲率與年降水量的計量單位不同,且數值差異較大,需對各原始數據進行均值化變換進行標準化無量綱處理(表2)。

2.3.3關聯系數和關聯度 根據關聯系數計算公式(2),可以求出各鼠種捕獲率與該地區年降水量的關聯系數(表4)。各鼠種捕獲率與年降水量的關聯系數在0.541 8~1.000 0。
將表4中的各關聯系數代入公式(3),計算出各鼠種的年捕獲率與年降水量的關聯度(表5)。關聯度的大小反映了年降水量對不同鼠種的作用大小,關聯度越大,則表示年降水量對該鼠種的影響越大。年降水量與群落中主要鼠種的關聯度大小順序為:三趾跳鼠>五趾跳鼠>小毛足鼠>阿拉善黃鼠>子午沙鼠>黑線倉鼠(表5)。因此,可以看出群落中跳鼠對降水量的反應較明顯,三趾跳鼠和五趾跳鼠的灰色關聯系數分別為0.855 6和0.818 9;子午沙鼠和黑線倉鼠對降水量的反應較弱,關聯系數僅為0.706 7和0.706 4。

表2 數據標準化處理

表3 年降水量與各鼠種捕獲率的絕對差值

表4 年降水量與各鼠種捕獲率的關聯系數
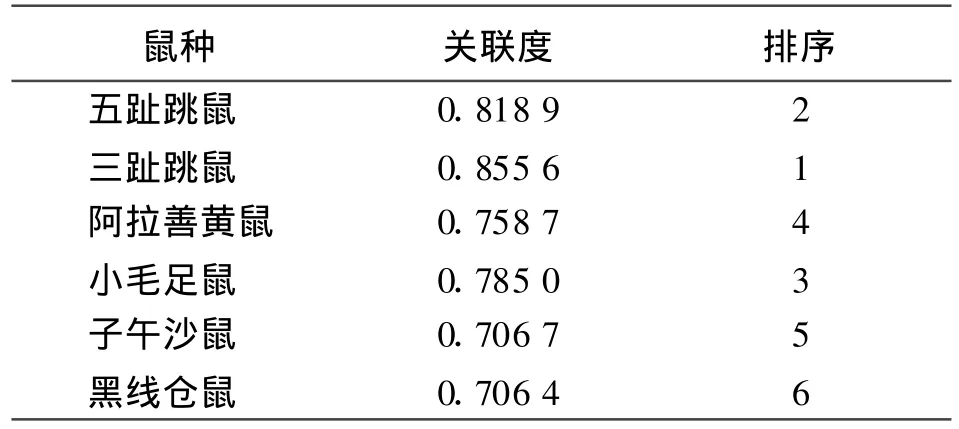
表5 群落中主要鼠種與全年降水量的關聯度
3 討論與結論
灰色系統理論是由我國學者鄧聚龍于1982年提出的[11]。近年來,該理論在工業、農業、生態和生物等許多領域得到廣泛應用[12-19],但國內外關于該方法在嚙齒動物研究上的應用鮮見報道。灰色關聯度分析是該理論中的重要分析方法,它的優點在于,對樣本量的多少和有無規律都同樣適用,不要求樣本服從某個典型概率分布,不要求各因素數據與系統特征數據之間呈線性關系,彌補了采用數理統計方法作系統分析所導致的缺憾。嚙齒動物種群數量與降水量之間的關系是復雜的,降水通過影響嚙齒動物的繁殖和食物資源等,影響其種群的大小。它們之間的關系是模糊的、灰色的,符合灰色理論中的灰色系統,因此灰色關聯度分析是揭示嚙齒動物與環境因子之間關系的一種有效方法。該方法的不足之處在于,關聯度分析法中只有正關聯度,無負關聯度,其范圍為0~1。而在數理統計學中的相關系數,有正、負和零,這符合生物界各種因素的相互影響有正、負相關性的客觀事實[13]。
本研究7年中在研究區共捕獲11個鼠種,選取了其中6個主要鼠種進行了分析。因為其他鼠種在各年度的捕獲數量較低或僅在個別年份中被捕獲,加以分析會影響關聯分析的準確性。例如,2008年長爪沙鼠的捕獲數量較大,為179只,而其他年份基本為0。長爪沙鼠在該地區主要生存在開墾農田生境中,2008年為長爪沙鼠的高峰年,可能由于其發生由農田向天然草地的擴散現象,造成該年天然草地中長爪沙鼠的數量明顯高于其他年份。因此,長爪沙鼠不是輪牧區中的主要組成物種,本研究中沒有對其進行灰色關聯度分析。
國內外關于嚙齒動物與降水量關系進行了大量研究,Ernest等[20]研究表明,美國Sevilleta地區的嚙齒動物種群大小和降水量是相關的。T riggs[5]發現小家鼠的種群數量與降水量變化有關。趙天飆等[6]研究表明大沙鼠(Rhombomys opimus)的種群數量與降水量顯著相關,其種群密度和當年降水量的變化基本一致 。本研究與以上研究結果相似,嚙齒動物種群數量與年降水量呈顯著正相關,說明降水量是影響嚙齒動物數量變動的重要因素之一。但是并不能忽視其他生態因子的作用,因為影響生物的因子是多方面的,多個因子共同作用決定生物的種群數量。因此,本研究中嚙齒動物數量的大小是由降水與其他生態因子共同作用決定的,有關各因子的作用大小還有待進一步研究。
近年來,生態學家們提出了資源脈沖理論[21],資源脈沖作用廣泛存在于生態系統中[22]。資源脈沖理論對于理解荒漠生態系統中嚙齒動物與降水量之間的關系是十分重要的。降水在荒漠生態系統中作為重要的脈沖資源,首先對植物種子的萌發、生長和繁殖產生作用,而植物又作為脈沖資源促進了嚙齒動物的生存和繁殖[20],決定了嚙齒動物的數量大小。因此,資源脈沖理論可以用來解釋本研究中嚙齒動物數量與年降水量呈顯著正相關的結果。本研究中2002-2007年,嚙齒動物數量與年降水量可以表現出正相關的變動趨勢,而2008年降水量較2007年降低,嚙齒動物的捕獲率卻明顯高于2007年。筆者認為滯后效應可能是造成該現象的原因,Letnic等[2]研究表明,在澳大利亞中部Simpson荒漠地區,嚙齒動物數量對于El Nio現象引起的降水變化表現出滯后效應。Ernest等[20]也發現嚙齒動物數量變化至少滯后于降水一個季節。因此,本研究中由于2007年的降水量較高,嚙齒動物數量對其反應表現出滯后效應,導致2008年嚙齒動物達到較高的數量。
不同嚙齒動物正常代謝過程中所需的水分平衡條件不同[23],導致其對降水量的反應也可能不同[24]。例如,內蒙古荒漠草原地區長爪沙鼠的種群密度與降水量是負相關的,而達烏爾鼠兔(Ochotona dauurica)種群增長率與降水量正相關[25-26]。本研究表明,群落中總的嚙齒動物種群數量與年降水量呈顯著正相關,但是各鼠種與年降水量的相關性并不一致。群落中主要鼠種與年降水量的關聯度順序為:三趾跳鼠>五趾跳鼠>小毛足鼠>阿拉善黃鼠>子午沙鼠>黑線倉鼠。子午沙鼠和小毛足鼠為群落中的優勢鼠種,它們與降水量的關系相對較弱,而跳鼠的種群大小對降水量的反應最明顯。可見,嚙齒動物對降水量變化的反應與其在群落中生態位的高低無關,可能與物種在長期進化中形成的生態適應機制有關,具體原因尚需進一步探討。
[1]Jaksic F M.Ecological effects of El Nio in terrestrial ecosystems of western South America[J].Ecography,2001,24(3):241-250.
[2]Letnic M,Tamayo B,Dickman C R.The responses of mammals to La Nia(El Nio southern oscillation)-associated rainfall,predation,and wildfire in central Australia[J].Journal ofMammalogy,2005,86(4):689-703.
[3]Meserve P L,Yunger J A,Gutié rrez J R,et al.Heterogeneous responses of small mammals to an El Nio southern oscillation event in northcentral semiarid Chile and the importance of the ecological scale[J].Journal of Mammalogy,1995,76(2):580-595.
[4]Jaksic F M,Lima M.M yths and facts on ratadas bamboo blooms,rainfall peaks and rodent outbreaks in South America[J].Austral Ecology,2003,28(3):237-251.
[5]Triggs G S.The population ecology of house mice(Musdomesticus)on the lsle of May,Scotland[J].Journal of Zoology,1991,225(3):449-468.
[6]趙天飆,楊持,周立志,等.大沙鼠種群密度與降水量的關系[J].獸類學報,2007,27(2):195-199.
[7]姜運良,盧浩泉,李玉春,等.山東陽谷縣黑線倉鼠種群數量預測預報[J].獸類學報,1994,14(3):195-201.
[8]Walsberg G E.Small mammals in hot deserts:some generalizations revisited[J].BioScience,2000,50(2):109-119.
[9]Lima M,M arquet P A,Jaksic F M.El Nio events,precipitation patterns,and rodent outbreaks are statistically associated in semiarid Chile[J].Ecography,1999,22(2):213-218.
[10]郭娜,郭科,吳金爐,等.灰色關聯度分析法在土地評價中的應用[J].成都理工大學學報(自然科學版),2007,34(6):626-629.
[11]鄧聚龍.灰色系統基本方法[M].武漢:華中理工大學出版社,1992.
[12]劉玉華,史紀安,賈志寬.氣候因子與苜蓿草產量的灰色關聯度分析[J].草業科學,2009,26(8):101-106.
[13]楊詩興.試論灰色系統關聯度分析法在生物科學研究中應用的合適性[J].家畜生態,2003,24(4):10-13.
[14]楊曌,張新全,李向林,等.應用灰色關聯度綜合評價17個不同秋眠級苜蓿的生產性能[J].草業學報,2009,18(5):67-72.
[15]蘇才旦,周雷.共和盆地溫性草原物候期草地生產力與氣候因素的灰色關聯度分析[J].草業科學,2009,26(5):38-43.
[16]劉玉成,杜道林,岳泉.緒云山森林次生演替中優勢種群的特性與生態因子的關聯度分析[J].植物生態學報,1994,18(3):283-289.
[17]蔡麗艷,石鳳翎,高翠萍,等.苜蓿雄性不育系雜交制種產量灰色關聯度分析[J].種子,2010,29(3):19-23.
[18]李云樂,喬玉輝,孫振鈞,等.不同土壤培肥措施下農田有機物分解的生態過程[J].生態學報,2006,26(6):1933-1939.
[19]馬姜明,李昆,張昌順.元謀干熱河谷蘇門答臘金合歡、新銀合歡人工林天然更新初步研究[J].應用生態學報,2006,17(8):1365-1369.
[20]Ernest S K M,Brown J H,Parmenter R R.Rodent,plant,and precipitation:spatial and temporal dynamics of consumers and resource[J].Oikos,2000,88(3):470-482.
[21]Holt R D.Theoretical perspectives on resource pulses[J].Ecology,2008,89(3):671-681.
[22]Orland M C,Kelt D A.Responses of a heteromyid rodent community to large-and small-scale resource pulses:diversity,abundance,and home-range dynamics[J].Journal of Mammalogy,2008,88(5):1280-1287.
[23]Tirado C,Corté s A,Bozinovic F.Water balance in two South American Phyllotis desert rodents,P.xanthopygus rupestrisandP.darwini darwini[J].Journal of Arid Environments,2008,72(5):664-670.
[24]Reed A W,Slade N A.Environmental correlates of survival and reproduction in old-field rodents[J].Journal of Mammalogy,2009,90(3):680-685.
[25]Wang G M,Zhong W Q.Mongolian gerbils and Daurian pikas responded differently to changes in precipitation in the Inner Mongolian grasslands[J].Journal of Arid Environments,2006,66(4):648-656.
[26]Liu W,Wan X,Zhong W Q.Population dynamics of the Mongolian gerbils:Seasonal patterns and interactions among density,reproduction and climate[J].Journal of Arid Environments,2007,68(3):389-397.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
