海洋交替單胞菌YTW-10的鑒定及抑菌活性分析
2011-03-15 10:12:20宿紅艷楊潤亞王曉杰林文杰
海洋科學 2011年7期
王 磊,宿紅艷,楊潤亞,王曉杰,林文杰
(魯東大學 生命科學學院,山東 煙臺 264025)
海洋占地球表面積的 71%,在遼闊的海洋中蘊藏著豐富的微生物資源。海洋微生物生活在海洋高鹽、高壓、低溫、低營養或無光照等特殊環境之中,這就使海洋微生物能產生陸棲微生物所不能產生的結構新穎、作用獨特的生物活性物質。因此,海洋微生物成為新藥篩選的重要資源,具有廣闊的發展前景,從海洋微生物中尋找生物活性物質已成為國內外研究的熱點[1-2]。海洋細菌是海洋微生物活性物質的一個重要來源,目前發現的具有潛在開發前景的主要包括芽孢桿菌屬(Bucillus)、交替單胞菌屬(Alteromonas)、假單胞菌屬(Pseudomonas)、假交替單胞菌屬(Pseudoalteromonas)、微球菌屬(Micrococcus)、腸桿菌屬(Enterubacterium)、黃桿菌屬(Flavobacterium)、著色菌屬(Chromtium)和欽氏菌屬(Chainia)[3]。研究表明,這些海洋微生物可產生抗生素等抗菌活性物質,對細菌、真菌、病毒等的生長有著明顯的拮抗作用[4-6]。
目前,病原真菌感染是植物病害的重要原因之一。不少由真菌引起的植物病害用傳統的化學農藥等方法尚不能有效地根治,同時化學農藥的大量使用容易帶來環境污染、土壤酸堿化及肥力下降等弊端。近年來人們利用對病原菌具有拮抗活性的微生物或者微生物的代謝物作為生物農藥進行生物防治成為一種既有效又環保的新途徑[7-8]。因此,高鹽環境中的嗜鹽菌作為一類新型的、極具應用前景的微生物資源,對其研究既具有理論意義,又有應用價值[9]。
作者從山東煙臺近海鹽場鹽池底泥中分離到一株具有抑菌活性的嗜鹽菌,通過對該菌的形態特征、培養特征、生理生化特性以及系統進化分析的研究,表明其屬于交替單胞菌。本實驗為豐富生物防治資源以及工業化生產抗真菌活性物質提供了科學依據,具有潛在藥物開發利用的價值。
1 材料和方法
1.1 材料
1.1.1 菌株
實驗菌株 YTW-10分離于山東煙臺近海鹽場鹽池底泥經抗菌活性的初篩對病原微生物具有明顯的抑制作用,本實驗室保存。
1.1.2 培養基
LB培養基:蛋白胨10 g,酵母提取物5 g,NaCl 10 g,蒸餾水定容至1 000 mL,調節pH至6.8~7.0。固體培養基則需加入瓊脂15~20 g。
PDA培養基:馬鈴薯200 g切成條,煮成黏液狀,過濾取汁,葡萄糖20 g,蛋白胨5 g,KH2PO41.0 g,蒸餾水定容至1 000 mL,瓊脂25 g。
2216培養基:酵母膏1 g,蛋白胨5 g,磷酸高鐵0.1 g,NaCl 30 g,蒸餾水定容至 1 000 mL,pH為7.2~7.4,瓊脂 20~25 g。
1.2 方法
1.2.1 菌落形態觀察
將經分離純化的菌株YTW-10接種于LB固體培養基平板上,28℃培養5 d,觀察其菌落大小、顏色、形態等特征。
1.2.2 生長條件的測定
最適鹽度實驗:供試菌株單菌落過夜震蕩培養,按照 1%接種量分別接種于 NaCl濃度為 1%、3%、5%、7%、10%、13%、15%的LB液體培養基中,28℃、150 r/min震蕩培養 12 h,測定培養液光密度值(A578)。重復3次實驗,取其平均值。
最適生長溫度實驗:將供試菌株單菌落過夜震蕩培養,按照1%接種量接種于含3% NaCl 的LB液體培養基中,分別置于4、20、30、37、45、55℃中150 r/min震蕩培養 36 h,測定培養液光密度值(A578)。重復3次實驗,取其平均值。
pH值對生長影響的測定:供試菌株單菌落過夜震蕩培養,按照1%接種量接種于含NaCl為3%,pH值分別為 4、5、6、7、8、9、10的 LB 培養液中 28℃、150 r/min震蕩培養 12 h,測定培養液光密度值(A578)。重復3次實驗,取其平均值。
1.2.3 生理生化實驗
參照《常見細菌系統鑒定手冊》[10]和《微生物分類學》[11]中的有關方法進行生理生化特征的鑒定。
1.2.4 16S rDNA序列測定與系統進化分析
基因組DNA的提取:YTW-10的基因組DAN提取采用寶生物工程(大連)有限公司的 TaKaRa Mini-BEST Bacterial Genomic DNA Extraction Kit Ver.2.0試劑盒。
16S rDNA 序列的 PCR 擴增和測序:以菌株YTW-10基因組DNA為模板,采用正向引物 P1:5’-AGA GTT TGA TCA TGG CTC AG - 3’和反向引物 P2:5’-AAG GAG GTG ATC CAG CCG CA-3’進行 PCR擴增。PCR 反應體系(25 μL):10×Buffer 2.5 μL,2.5 mmol/ L MgCl22.0 μL,2.5 mmol/ L dNTP 2.0 μL,TaqDNA 聚合酶 1 U,10 μmol/L 引物各 2.0 μL,5~10 ng基因組DNA,雙蒸水補齊體積。擴增條件:94 ℃預變性10 min,94 ℃變性 1 min,55 ℃退火1 min,72 ℃延伸2 min,循環35次,72 ℃后延伸5 min。PCR 擴增產物經Elute milk小量膠回收試劑盒(上海飛捷生物技術有限公司)純化后,送上海生物工程有限公司測序。
系統進化樹的構建和分析:將測定的16S rDNA序列在GenBank數據庫中進行BLASTN相似性搜索,尋找同源序列。將分值較高的序列同本研究獲得的相應序列采用DNASTAR軟件包中的MegAlign程序進行多序列比對,并采用鄰接法(Neighbor-Joining)構建系統進化樹[12]。
1.2.5 抑菌活性實驗
指示菌株:鰻弧菌(Vibrio anguillarum)、溶藻弧菌(Vibrio alginolyticus)、副溶血弧菌(Vibrio parahaemolyticus)等水產生物致病細菌購于上海水產大學水產動物病原庫; 玉米彎胞病菌(Curvlaria lunata)、西瓜枯萎病菌(Fusarium ozysporum)和小麥赤霉病菌(Fusarium graminearumSchw)等植物致病真菌由西北農林科技大學植物病理實驗室提供。
指示菌培養基:水產生物致病菌采用2216培養基,植物致病真菌采用葡萄糖土豆(PDA)培養基。
供試菌對病原細菌抑菌活性實驗:將培養至對數增長期的病原細菌培養液150 μL均勻涂布在2216固體培養基平皿上,然后在培養基中央上打孔,加入50 μL經搖床培養24 h的YTW-10培養液。在30℃的溫箱中培養72 h后,測量抑菌圈橫向和縱向直徑,取平均值。
供試菌對病原真菌抑菌活性實驗:將平板培養的指示真菌用打孔器制成圓形菌苔接種到PDA培養基中央,然后在真菌周圍一定范圍內均勻打 4個孔,分別加入50 μL經搖床培養24 h的YTW-10培養液,同時以不加 YTW-10培養液的真菌平板作為對照。在30 ℃的溫箱中培養72 h,分別測量對照皿和供試皿中的真菌菌落直徑,二者的差值即為抑菌直徑[13]。
2 結果
2.1 菌落形態觀察
菌株YTW-10在LB固體培養基上菌落形狀為圓形,表面光滑,扁平凸起,邊緣整齊,菌落大小(d/mm)為 1~2,表面濕潤,白色,不透明。革蘭氏染色陰性。具備《常見細菌系統鑒定手冊》中交替單胞菌屬的特征。
2.2 生長條件測定
將YTW-10接種在不同NaCl濃度的LB液體培養基中培養,繪制其在不同鹽濃度下的生長曲線。由圖 1可以看出YTW-10可在1%~15 % NaCl濃度的LB培養基中生長,在NaCl濃度為7%左右生長最好,當 NaCl濃度在 10%以上時其生長量急劇下降,到15%時幾乎不生長,因此我們將其定義為中度嗜鹽菌。
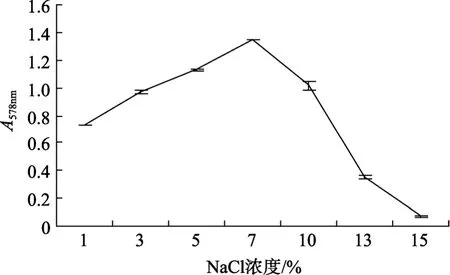
圖1 YTW-10在不同 NaCl濃度的 LB 培養基中的生長情況Fig.1 Growth of YTW-10 in LB medium with different concentrations of NaCl
從圖 2可以看出YTW-10菌株生長的溫度范圍較為廣泛,從4 ℃到55 ℃都有不同程度的生長,在30~45 ℃之間生長較好,在 37℃時生長最快;YTW-10的pH值生長范圍在4~10之間,在4和10時幾乎不生長,在pH為8時生長速度最快(圖3)。
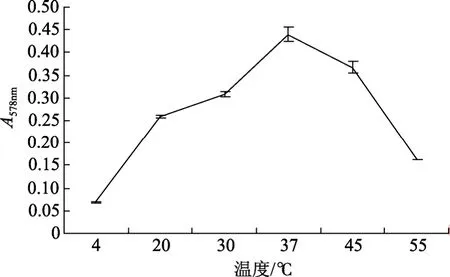
圖2 YTW-10在不同溫度下生長的情況Fig.2 Growth of YTW-10 at different temperatures
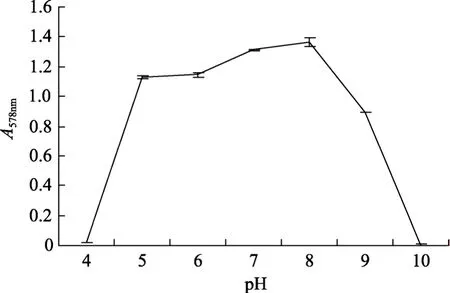
圖3 pH值對YTW-10生長的影響Fig.3 Effects of pH on YTW-10 growth
2.3 理化性質實驗
菌株 YTW-10 的生理生化特征如表1所示,可以利用木糖、麥芽糖、蔗糖、葡萄糖、甘露糖為唯一碳源生長,但不能利用核糖和阿拉伯糖作為唯一碳源生長; 能以水解明膠作為唯一氮源生長,不能利用酪蛋白為唯一氮源生長; 能產硫化氫,葡萄糖氧化產酸(M.R 反應為陽性),V.P反應為陰性; 其過氧化氫酶、氧化酶、卵磷脂酶為陽性,反硝化、精氨酸脫羧、苯丙氨酸脫氨、淀粉水解等實驗均為陰性。
初步鑒定結果表明,YTW-10的形態和生理生化特性與交替單胞菌屬較為一致,YTW-10與交替單胞菌的兩個確定種的生理鑒定結果的比較見表1。
2.4 基于16S r DNA序列的系統進化分析
為進一步鑒定 YTW-10,在理化分析的基礎上,對 YTW-10進行 16S rDNA序列分析。通過對YTW-10的16S rDNA序列擴增獲得了1 494 bp的序列,并提交到GenBank中(登陸號為FJ171338)。將該16S rDNA序列在NCBI數據庫中進行Blastn比對,結果表明在前50個同源性較高的序列中已確定的屬名均為交替單胞菌屬,同源性為 99 %,但全部是未定種。其中比對分值最高的是Alteromonassp.CF14-3和Alteromonassp.CF11-5的 16S rDNA 序列。與YTW-10同源性最高的已確定種是Alteromonas macleodii,同源性為 97%。選取與 YTW-10同源性較高的代表菌株進行系統進化分析,以交替單胞菌科(Alteromonadaceae)中的另一個屬假交替單胞菌屬(Pseudoalteromonas)為外類群構建系統進化樹(圖4)。系統進化樹表明YTW-10與所有的Alteromonas聚在一起,其中YTW-10與Alteromonassp.CF14-3的親緣關系最近。結合前期完成的培養特征和生理生化特征等實驗的結果,我們最終確定 YTW-10為交替單胞菌屬Alteromonas細菌。
2.5 抑菌實驗
以水產生物致病菌鰻弧菌、溶藻弧菌和副溶血弧菌以及植物致病真菌玉米彎胞病菌、 西瓜枯萎病菌和小麥赤霉病菌為敏感指示菌,進行拮抗對峙實驗,抑菌譜結果見表2。
實驗結果表明 YTW-10對水產生物致病細菌的拮抗作用不明顯,但對植物致病真菌具有較強的抑菌活性。圖5所示是YTW-10對不同植物致病真菌生長的抑制效果,結果表明沒有接種 YTW-10的植物致病真菌生長速度較快,長勢好; 而接種YTW-10的生長速度緩慢很多,長勢較差。因此,菌株YTW-10對小麥赤霉病菌、西瓜枯萎病菌和玉米彎胞病菌都有明顯的抑制效果。
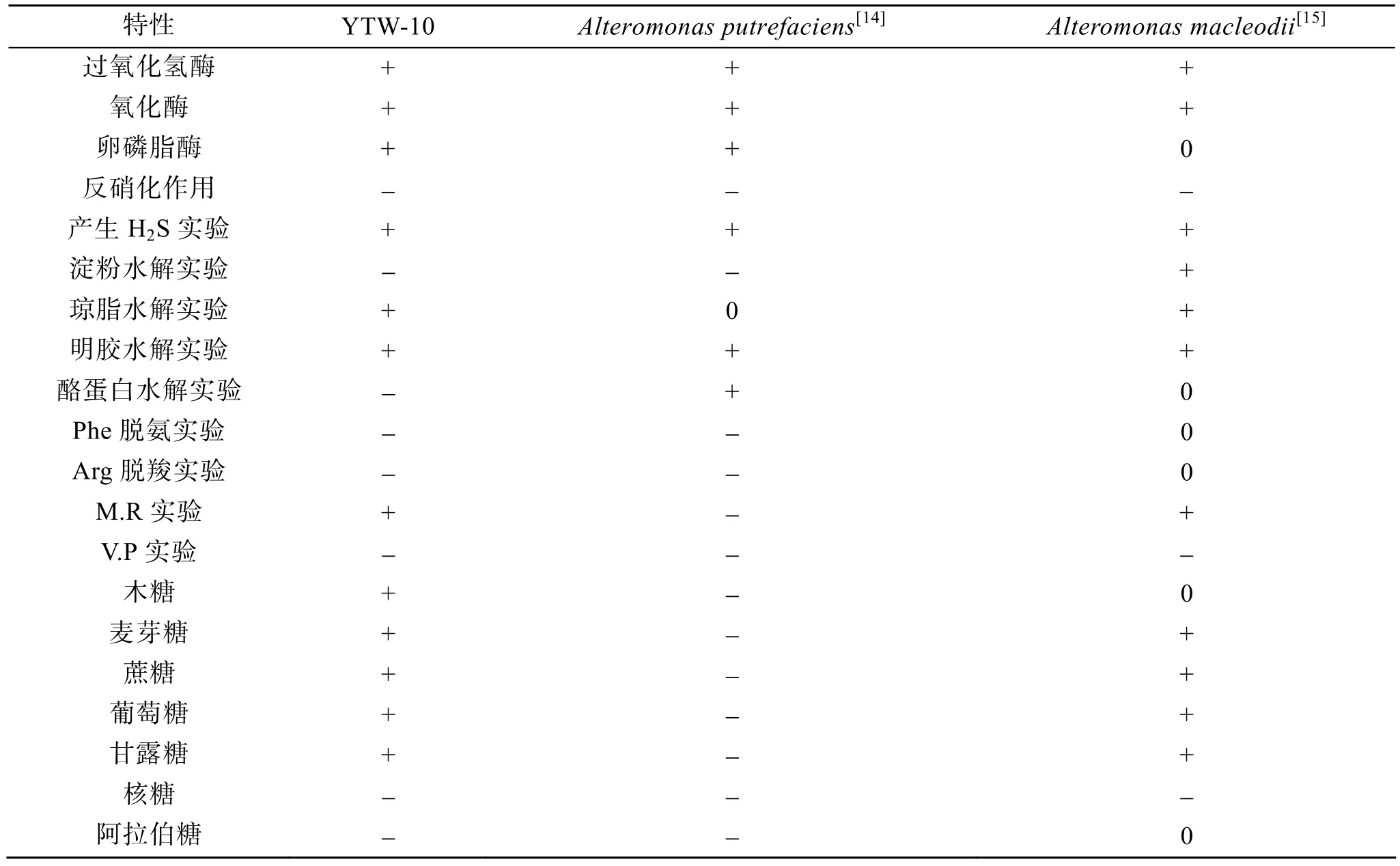
表1 菌株YTW-10與相近菌株生理生化特征比較Tab.1 Comparison of physiological and biochemical characteristics between YTW-10 and the related strains
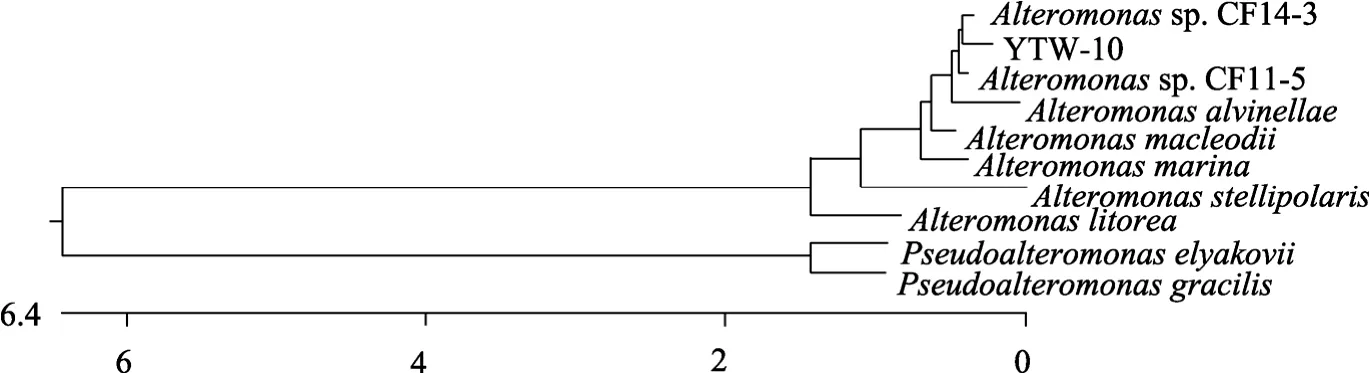
圖4 YTW-10菌株的16S rDNA 序列的系統進化樹Fig.4 Phylogentic tree of YTW-10 based on 16S rDNA sequences
3 討論
陸生微生物發酵產物長期以來一直是醫藥業和工農業的重要來源,然而由于該領域研究的深入,近年來從陸生微生物中尋找新的資源難度越來越大。另外傳統抗生素的濫用,使得耐藥病原菌不斷出現[16]。由于海洋微生物獨特的生存環境,海洋微生物中常蘊含著陸生生物所不具備的活性物質[17]。 因此,從海洋微生物的次級代謝產物中尋找開發有別于陸棲微生物產生的生理活性物質已成為當前微生物學研究的一個重要方向。已有的研究表明海洋細菌的代謝產物種類繁多,結構新穎多樣,存在著豐富的抗菌資源[18]。我國海岸線長,海域遼闊,蘊含著豐富的海洋微生物資源,需要加強這方面的研究和開發工作。

表2 YTW-10的抑菌譜Tab.2 Inhibition spectra of YTW-10
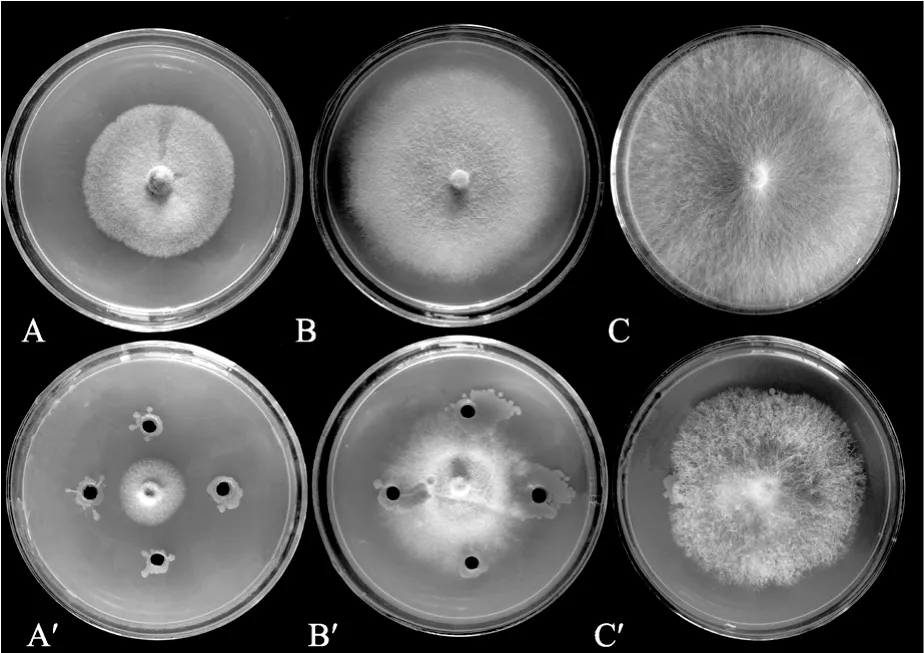
圖5 菌株YTW-10對病原真菌的抑制Fig.5 Antibacterial test
作者從煙臺海域分離到一株對植物致病真菌具有明顯拮抗作用的菌株YTW-10,根據YTW-10的形態學特征和生理生化鑒定,YTW-10與交替單胞菌較為接近。基于16S rDNA序列的分子生物學鑒定顯示,該菌在基因庫中與未確定種名的Alteromonassp.CF14-3最為接近,相似度達99%; 與YTW-10最接近的確定種是Alteromonas macleodii,同源性為97%。應用 16S rDNA 序列分析法能快速、準確地對微生物進行分類鑒定,在環境微生物多樣性研究、菌種鑒定及微生物資源的開發中得到了廣泛的應用,目前隨著分子分類的理論和方法的日趨成熟以及數據庫的日趨完善,它已經逐步成為微生物資源調查和環境生態研究的一種強有力的工具。一般認為,16S rDNA 序列同源性小于 98%,可以認為屬于不同的種[19]。因此,YTW-10有可能是交替單胞菌屬的潛在新種,值得進一步研究。
交替單胞菌屬的菌能夠產生多種新穎活性物質,易實現工業化的生產,具有廣闊的開發前景[20]。杜宗軍等[21]從青島近海海水中分離得到的交替單胞菌Alteromonas addita具有降解瓊膠活性。王海麗等[22]利用取自南極的交替單胞菌Alteromonas stellipolaris提取外膜蛋白和脂多糖,研究它們對黑鯛非特異性免疫功能的影響,發現可以起到免疫增強劑的作用,能增強黑鯛血清中的抗菌活力、溶菌酶的活力和酚氧化酶活力,顯著提高黑鯛的白細胞吞噬活性。鄭立等[23]對從海洋生物、海水和海泥中分離的海洋細菌進行抗菌活性研究,發現多種交替單胞菌對枯草芽孢桿菌、金黃色葡萄球菌、根瘤土壤桿菌具有不同效果的抑菌作用。目前,國內對交替單胞菌研究相對較少,特別是在其產生抗病原真菌活性物質方面的研究尚屬空白。我們首次對從煙臺海域篩選分離得到的 YTW-10開展針對抗水產致病菌和植物病原菌的活性研究,發現其具有廣譜的抗植物病原真菌的作用,為豐富生物防治資源以及工業化生產抗真菌活性物質提供了科學依據。
[1]Frette L,Johnsen K,Jorgensen N, et al.Functional characteristics of culturable bacterioplankton from marine and estuarine environments[J].Int Microbiol,2004,7(3):219-227.
[2]林白雪,黃志強,謝聯輝.海洋細菌活性物質的研究進展[J].微生物學報,2005,45(4):657-660.
[3]鄧祖軍,曹理想,譚紅銘,等.一株秋茄內生真菌(MF59)對植物病原真菌拮抗活性的研究[J].科技論壇,2007,7:86-89.
[4]姜健,楊寶靈,元起,劉松梅,等.具有抑菌活性的海洋細菌的分離與鑒定[J].生物技術,2004,6:28-30.
[5]林敏,寧喜斌.具抑菌活性海洋微生物的篩選[J].微生物學雜志,2005,5:23-25.
[6]詹萍,梁靜娟,龐宗文,等.假交替單胞菌JIV_49產抗真菌活性物質的發酵條件研究[J].生物技術,2007,1:69-72.
[7]Sun Y,Tian L,Huang Y F,et al.A new cyclotetrapeptide from marine fungusTrichoderma reesei[J].Pharmazie,2006,61(9):809-810.
[8]石瑛,田黎,王婧,等.海洋放線菌Micromonosporasp.與細菌Oceanospirillumsp.發酵液中化學成分的研究[J].中國海洋藥物,2006,25(1):6-10.
[9]張銳,曾潤穎.極端微生物產堿性蛋白酶菌株的篩選及發酵條件研究[J].微生物學通報.2001,28(4):5-8.
[10]東秀珠,蔡妙英.常見細菌系統鑒定手冊[M].北京:科學出版社,2001:99-121.
[11]沈萍,范秀容,李光武.微生物學實驗[M].北京:高等教育出版社,2004:215-221.
[12]Burland,T G.DNASTAR's Lasergene sequence analysis software[J].Methods in Molecular Biology,2000,132:71-91.
[13]曹理想,田新莉,周世寧.香蕉內生真菌、放線菌類群分析[J].中山大學學報(自然科學版),2003,42(2):70-73.
[14]鮮海軍.對兩株具有多種染料脫色能力的腐敗交替單胞菌(Alteromonas putrefaciens)的研究[J].應用與環境生物學報,2002,8(3):294-297.
[15]Han B Q,Dai J X,Wang H.Isolation and identification of algina-degrading bacteria and formation of alginase[J].Acta Oceanologica Sinica,1999,l18(4):555-561.
[16]程福亮,韓文瑜,雷連成,等.33株臨床分離的藥敏試驗分析[J].動物醫學進展,2007,28(2):15-18.
[17]李越中,陳琦.海洋微生物資源及其產生生物活性代謝產物的研究[J].生物工程進展,2000,20(5):28-31.
[18]李大志,韓寶芹.海洋細菌活性代謝產物的研究進展[J].中國海洋藥物雜志.2006,25(5):51-55.
[19]Fry N K,Warwick S,Saunders N A,et al.The use of 16S ribosomal RNA analyses to investigate the phylogeny of the family Legionellaceae [J].J Gen Microbiol,1991,137(5):1215-1222.
[20]席宇,朱大恒,劉紅濤,等.假交替單胞菌及其胞外活性物質研究進展[J].微生物學通報,2005,32(3):108-112.
[21]杜宗軍,趙苑,李美菊,等.青島近海瓊膠降解細菌的篩選和多樣性分析[J].中國海洋大學學報,2007,37(2):277-282.
[22]王海麗.楊季芳.南極超微細菌 ANT52(Alteromonas stellipolaris)外膜蛋白和脂多糖的提取物對黑鯛的免疫活性研究[J].海洋與湖沼2009,40(2):242-248.
[23]鄭立,林偉,嚴小軍,等.海洋細菌抗菌和細胞毒活性的初步研究[J].應用生態學報,2004,15(9):1633-1636.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
