長江中下游地區湖北釘螺線粒體CO I基因遺傳變異研究
2011-03-14 06:15:04李洪軍曲國立邢云天李幼子魏劍英戴建榮梁幼生
海洋科學 2011年10期
關鍵詞:研究
李洪軍,汪 偉,張 偉,曲國立,邢云天,李幼子,魏劍英,戴建榮,梁幼生
(1.江蘇省血吸蟲病防治研究所,江蘇 無錫 214064; 2.衛生部 寄生蟲病預防與控制技術重點實驗室,江蘇無錫 214064)
血吸蟲病是一種嚴重危害人類健康、影響社會經濟發展的“被忽略”的熱帶病(Neglected Tropical Diseases)[1],全球有76個國家和地區流行血吸蟲病,感染者超過2億,近8億人口面臨感染威脅[2]。在我國,日本血吸蟲病仍然是重要的公共衛生問題[3-5]。截至2010年底,全國仍有血吸蟲病患者約51.6萬[6]。經過多年積極防治,我國的血吸蟲病流行區主要被壓縮在長江中、下游沿岸5省(湖南、湖北、江西、安徽和江蘇)的江湖洲灘地區[7-10]。釘螺是日本血吸蟲的唯一中間宿主,是構成血吸蟲病傳播的不可缺失環節[11]。研究表明,凡有血吸蟲病流行的地區必有釘螺孳生[12]。控制與消滅釘螺是阻斷血吸蟲病傳播的有效措施之一[13-14]。
既往已有采用現代分子生物學技術對中國大陸釘螺系統進化進行研究的報道[15-18],亦有學者對湖北省廟河地區和長江三峽庫區釘螺線粒體基因遺傳變異進行了探討[19-20]。本研究通過對長江中、下游血吸蟲病流行區江蘇、安徽、江西、浙江和湖北 5省8個地理株湖北釘螺線粒體DNA細胞色素C氧化酶亞單位1(CO I)基因進行遺傳多態性分析,探討其核酸特異性,為進行長江中、下游地區釘螺遺傳變異研究、建立DNA 指紋檢測平臺及釘螺分子鑒定提供參考依據。
1 材料與方法
1.1 材料
1.1.1 釘螺樣本采集
分別采自長江中、下游地區的江蘇、湖北、安徽、江西、浙江4個省的8個流行區現場釘螺,釘螺采集地分別為江蘇丹徒和邗江、湖北武昌、安徽安慶和貴池、江西彭澤和余干、浙江平湖。所有釘螺均為肋殼釘螺,系湖北釘螺指名亞種(Oncomelania hupensis hupensis)。在實驗室內適應性飼養后,采用群體逸蚴法鑒定釘螺是否感染血吸蟲,選擇陰性成螺備用。
1.1.2 主要儀器與試劑
PCR儀為德國analyticjena公司產品,臺式離心機為德國Eppendorf 公司產品; 基因組DNA抽提試劑盒購自 promega公司,Taq DNA 聚合酶購自promega公司。
1.2 方法
1.2.1 釘螺基因組DNA的提取
選擇每一地理株釘螺3~5只,徹底洗凈后,去除螺殼及內臟腸管組織,將剩余組織進行混合,移入 EP管中,加入蛋白激酶 K,混勻,55 ℃水浴過夜。用基因組 DNA抽提試劑盒提取釘螺基因組DNA,并在核酸蛋白分析儀上測定 DNA濃度和純度。
1.2.2 PCR擴增及產物鑒定、測序
上游引物 P1:5′-GGTCAACAAATCATAAAGATATTGG-3′,下游引物 P2:5′ -TAAACTTCAGGGTGACCAAAAAAYCA-3′,由上海生物工程有限公司合成。50 μL 反應體系為:10×reaction buffer 5.0 μL,25mmol/L MgCl25.0 μL,dNTP 2.0 μL,上、下游引物各 2.0 μL,模板 DNA 1.0 μL,Taq DNA聚合酶 1.0 μL,用ddH2O補足反應體系。反應循環條件為:94 ℃預變性10 min,94 ℃變性1 min,55 ℃退火90 s(COI),72 ℃延伸1 min,共35個循環,末次循環72℃延伸10 min。擴增產物經1.5%瓊脂糖凝膠電泳鑒定,并將擴增產物送上海英駿生物技術公司測序。
1.2.3 CO I 基因序列分析
將測序結果先用ClustalX 1.81軟件進行多序列比對,再在 MEGA 4.0軟件中的 Kimura 雙參數法(Kimura 2-Parameter)計算遺傳距離,分別用鄰接法(NJ)和最大簡約法(MP) 構建系統發生樹,Bootstrap進行檢驗。
2 結果
2.1 CO I 基因PCR 擴增結果
以 8個不同地理株湖北釘螺基因組 DNA 為模板,PCR 特異性擴增線粒體DNA CO I 基因,均在長度約700 bp 附近均擴增出一目的條帶,與預期目的片段大小一致。
2.2 CO I基因序列分析
經過 ClustalX 1.81 軟件比對,刪除多變區后,得到701個同源位點,其中有保守位點663個,可變位點38個,簡約信息位點26個。A、T、C和G的堿基平均含量分別為22.8%、37.6%、18.8%、20.8%,A+T平均含量為 60.4%,C+G平均含量為 39.6%,A/T含量明顯偏高。表1給出了基于Kimura雙參數模型序列分歧矩陣。最終得出所有序列間的平均遺傳距離為0.025(表1)。
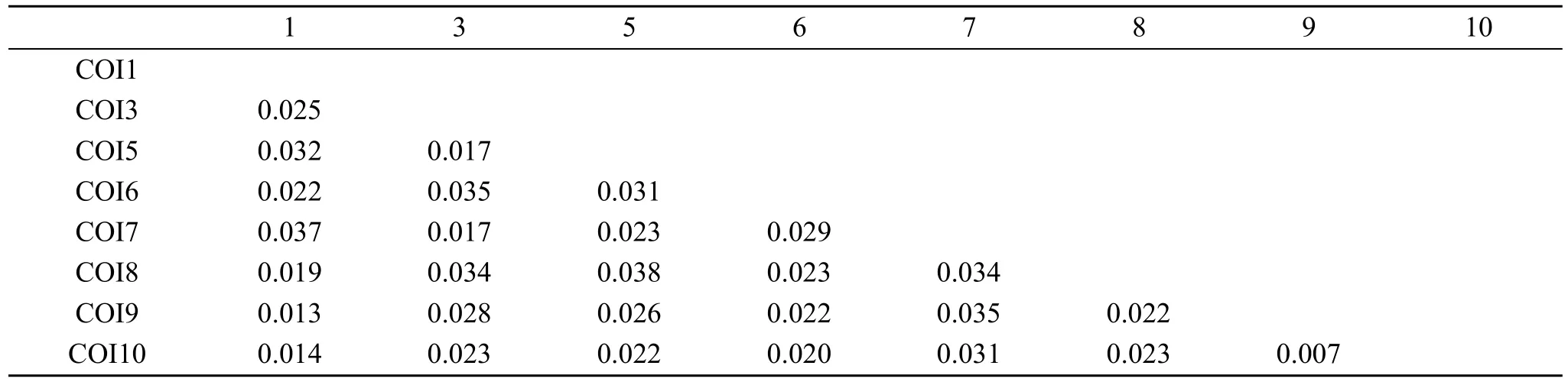
表1 本研究中不同地理株間的遺傳距離(基于Kimura雙參數模型)Tab.1 The genetic distances based on the formula of Kimura 2 parameters among snails from different areas in this study
2.3 基于CO I基因序列的分子系統樹
本研究采用鄰接法和最大簡約法共構建了 2種分子系統樹:NJ樹和 MP樹。NJ樹中的分支檢驗置信值和后驗值低于 50%的則未顯示。NJ進化樹顯示,江蘇丹徒、湖北武昌、江西余干及安徽安慶和貴池5個地理株的釘螺形成一個分支,而浙江平湖、江蘇邗江和江西彭澤3個地理株釘螺形成另一個分支; 但兩種方法構建的系統發生樹存在差異。在內部分支檢驗置信值在大于 80%時很相近(圖 1、圖 2)。

圖1 基于CO I基因部分序列構建的NJ樹(各分支上數字為1 000次內部分支檢驗置信值)Fig.1 The NJ tree based on CO I gene sequence data(Numbers on each node correspond to their bootstrap values for 1000 replicates.)
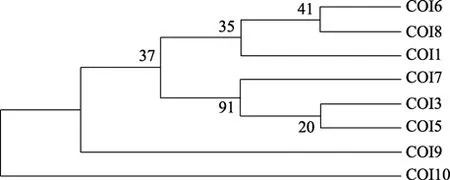
圖2 基于CO I基因部分序列構建的MP樹各分支上的數字為bootstrap1 000個循環的自舉檢驗值Fig.2 The MP tree based on partial of CO I gene sequences data with confident values of Bootstrap 1 000 indicated above each branch
3 討論
隨著分子生物學技術的迅速發展,線粒體DNA、核糖體DNA和RNA、微衛星DNA、單核苷酸多態性(SNP)和限制性片段長度多態性(RFLP)、隨機擴增多態性(RAPD)等多種分子遺傳研究方法和遺傳標記的出現,為湖北釘螺遺傳多態性及其分子系統學的研究提供了更豐富的手段[21-22]。線粒體DNA作為一種核外遺傳物質,具有以母性遺傳為主,進化速率快,較易發生突變,其突變頻率約為核基因組的 5~10 倍,且基因重組率極低,同一個體組織中的線粒體 DNA 具有一致性等特點,使它成為生物進化過程中譜系發生和遷移流動的有效遺傳標記,已經廣泛應用于寄生蟲屬、種及種群水平相互關系的研究[19,23-26]。
本研究通過對長江中、下游地區 8個不同地理株湖北釘螺線粒體 DNA CO I 基因進行序列分析,從分子水平上探明其種群內的遺傳差異。研究發現,不同地域株湖北釘螺遺傳差異顯著,平均遺傳距離為 0.025。進化樹顯示,江蘇丹徒、湖北武昌、江西余干及安徽安慶和貴池 5個地理株釘螺形成一個分支,而浙江平湖、江蘇邗江和江西彭澤3個地域釘螺形成另一個分支。這可能與長江中、下游地區形成的天然地理隔離和生態環境差異有關,支持 Davis等[27]對湖北釘螺的分類結果。Wilke等[17]對釘螺cox1基因序列研究發現,長江中下游地區光殼和肋殼釘螺之間遺傳變異很小,同屬指名亞種。石朝輝等[28]發現同一地區光殼與肋殼釘螺 cox1 基因序列之間有較高的同源性。本實驗結果表明,長江中、下游地區湖北釘螺種群內存在較高的核苷酸多態性,遺傳差異較顯著,且兩種方法構建的進化樹存在差異。由于線粒體涉及絕大多數真核生物的呼吸代謝,線粒體DNA 含有12 或13 種呼吸鏈酶原復合物中重要蛋白的編碼基因,具有豐富的遺傳特征,在生物進化過程中受到的選擇壓力較大。因此,線粒體 DNA序列的進化要快于細胞核基因[29]。這一遺傳差異的原因及其會否導致螺宿主對日本血吸蟲易感性差異有待于進一步研究。
本研究采用線粒體DNA CO I基因分子標記獲得了長江中、下游血吸蟲病流行區湖北釘螺種群CO I基因序列差異,為進一步構建中國大陸湖北釘螺遺傳多態性數據庫和DNA 指紋檢測平臺奠定了基礎。建立的分子技術可對現場采集到的釘螺進行地理種株和親緣關系鑒定。
[1]O’Connell D.Neglected Diseases[J].Nature,2007,449:157-182.
[2]Steinmann P,Keiser J,Bos R,et al.Schistosomiasis and water resources development:systematic review,meta-analysis,and estimates of people at risk[J].Lancet Infect Dis,2006,6:411-425.
[3]周曉農,姜慶五,汪天平,等.我國血吸蟲病防治研究現狀與發展戰略思考[J].中國血吸蟲病防治雜志,2004,17(1):1-3.
[4]Zhou X N,Guo J G,Wu X H,et al.Epidemiology of schistosomiasis in the People’s Republic of China,2004[J].Emerg Infect Dis,2007,13:1470-1476.
[5]Wang L D,Utzinger J,Zhou X N.Schistosomiasis control:experiences and lessons from China[J].Lancet,2008,372:1793-1795.
[6]郝陽,鄭浩,朱蓉,等.2009年全國血吸蟲病疫情通報[J].中國血吸蟲病防治雜志,2010,22(6):521-527.
[7]Li S Z,Luz A,Wang X H,et al.Schistosomiasis in China:acute infections during 2005-2008[J].Chin Med J (Engl),2009,122:1009-1014.
[8]郝陽,易冬華,張險峰,等.2008年全國血吸蟲病疫情控制考核評估報告[J].中國血吸蟲病防治雜志,2009,21(6):457-463.
[9]Zhao G M,Zhao Q,Jiang QW,et al.Surveillance for schistosomiasis japonica in China from 2000 to 2003[J].Acta Trop,2005,96:288-295.
[10]朱蓉,黨輝,張利娟,等.2005-2008年全國血吸蟲病疫情監測[J].中國血吸蟲病防治雜志,2009,21(5):358-362.
[11]中華人民共和國衛生部疾病控制司.血吸蟲病防治手冊[M].3版.上海:上海科學技術出版社,2000:23.
[12]袁鴻昌,張紹基,姜慶五.血吸蟲病防治理論與實踐[M].上海:復旦大學出版社,2003:3-4.
[13]郭家鋼.中國血吸蟲病綜合治理的歷史與現狀[J].中華預防醫學雜志,2006,40(4):225-228.
[14]Yuan Y,Xu X J,Dong H F,et al.Transmission control of schistosmiasis japonica:implementation and evaluation of different snail control interventions[J].Acta Trop,2005,96:191-197.
[15]劉月英,樓子康,王耀先,等.釘螺亞種分化[J].動物分類學報,1981,6(3):253-266.
[16]周曉農,孫樂平,洪青標,等.中國大陸釘螺種群遺傳學研究 I 種群遺傳差異[J].中國血吸蟲病防治雜志,1995,7(2):67-71.
[17]Wilker T,Davis G M,Cui E C,et al.Oncomelania hupensis(Gastropoda:Rissooidea) in eastern China:molecular phylogeny,population structure,and ecology[J].Acta Trop,2000,77:215-227.
[18]周藝彪,趙根明,韋建國,等.湖北釘螺種群內AFLP分子標記遺傳變異分析[J].中國寄生蟲學與寄生蟲病雜志,2006,24(1):27-31.
[19]Shi C H,Wilke T,Davis G M,et al.Population genetics,microphylogeography,ecology and susceptibility to schistosome infection of ChineseOncomelania hupensis hupensis(Gastropoda:Rissooidea:Pomatiopsidae)in the Miao River System[J].Malacologia,2002,44:333-338.
[20]陳琳,張錫林,何諧,等.三峽庫區上、下游血吸蟲病流行區釘螺線粒體cox1基因遺傳變異研究[J].熱帶醫學雜志,2010,10(1):5-8.
[21]周藝彪,姜慶五,趙根明.湖北釘螺遺傳多樣性及其分子系統學研究進展[J].中國血吸蟲病防治雜志,2005,17(5):391-396.
[22]孫恩濤,張錫林.湖北釘螺遺傳標記研究進展[J].中國血吸蟲病防治雜志,2010,22(1):90-95.
[23]Moro P L,Nakao M,Ito A,et al.Molecular identification ofEchinococcusisolates from Peru[J].Parasitol Int,2009,58:184-186.
[24]Nguyen T G,Van De N,Vercruysse J,et al.Genotypic characterization and species identification ofFasciolaspp.with implications regarding the isolates infecting goats in Vietnam[J].Exp Parasitol,2009,123:354-361.
[25]Jeon H K,Kim K H,Huh S,et al.Morphologic and genetic identification of Diphyllobothrium nihonkaiense in Korea[J].Korean J Parasitol,2009,47:369-375.
[26]Semyenova S K,Morozova E V,Chrisanfova G G,et al.Genetic differentiation in eastern European and western Asian populations of the liver fluke,Fasciola hepatica,as revealed by mitochondrial nad1 and cox1 genes[J].J Parasitol,2006,92:525-530.
[27]Davis G M,Wilke T,Zhang Yi,et al.Snails-Schistosoma,paragonimus interactions in China:population ecology,genetic diversity,coevolution and emerging diseases[J].Malacologia,1999,41:355-377.
[28]石朝輝,邱持平,夏明儀,等.湖北省廟河地區釘螺細胞色素C 氧化酶1 基因差異的研究[J].中國寄生蟲學與寄生蟲病雜志,2001,19(1):41-44.
[29]Le T H,Blair D,McManus D P.Mitochondrial genomes of parasitic flatworms [J].Trends Parasitol,2002,18:206-213.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
