人類博卡病毒HBoV1基因組克隆及啟動子活性分析
2011-02-10 01:20:32李京京孫彬歐陽錦鳳陳瑩韓虎劉凱于李毅
生物工程學報 2011年6期
關鍵詞:檢測
李京京,孫彬,歐陽錦鳳,陳瑩,韓虎,劉凱于,李毅
1 華中師范大學生命科學學院,武漢 430079
2 湖北師范學院生命科學學院,黃石 435002
呼吸道病毒是引發嬰幼兒呼吸道疾病的主要病原體,目前仍有許多新病毒被不斷發現。2005年,Allander等[1]采用隨機 PCR擴增及生物信息學分析,從下呼吸道感染患兒標本中檢測到一種新的病毒,命名為人博卡病毒 (Human bocavirus,HBoV)。隨后,許多國家相繼報道發現HBoV感染,且在患有腸胃炎的小兒糞便中也鑒定出人博卡病毒[2]。HBoV主要感染 2歲以下嬰幼兒,臨床癥狀主要表現為咳嗽、發熱常伴有重癥哮喘[3],伴隨其他呼吸道病毒共感染[4-5]。基因組序列分析表明,該病毒與牛細小病毒 (BPV) 及犬微小病毒 (MVC) 在系統發育上接近,將其歸屬于細小病毒亞科博卡病毒屬。最近,在胃腸道疾病患者的糞便中又發現了 3種不同基因型的人博卡病毒 HBoV2、HBoV3和HBoV4[6-8],證明博卡病毒不僅是小兒呼吸道感染的病原體,還會引發包括成人的病毒性腸胃炎疾病。
HBoV為單鏈、線狀 DNA病毒,全長約5 200 nt,由位于基因組左端的唯一啟動子啟動基因轉錄[9],該特征與紅細胞病毒屬、貂阿留申病毒屬的許多病毒相同,如人細小病毒B19基因組由唯一啟動子 P6調控 12種轉錄子合成[10-12]。2個主要開放閱讀框 (ORF) 分別編碼非結構蛋白 NS1和衣殼蛋白VP1/VP2,一個次要ORF編碼NP1,其功能未知。由于博卡病毒屬其他2個成員基因組兩端都具有ITR序列,該結構對病毒DNA復制起著重要作用[13],推測博卡病毒也具有ITR序列,但目前還未得到相關序列。Dijkman等首次報道,在HBoV1感染的假分層人氣管上皮細胞中檢測到 6個轉錄產物,且通過real-time PCR檢測到病毒復制[9]。隨后,Chen等報道了更完整的HBoV1轉錄圖譜,轉錄子(Transcripts) 均由位于基因組左端的P3啟動子啟動轉錄,轉錄圖譜特征與博卡病毒屬其他 2位成員類似[14]。細小病毒的敏感宿主細胞范圍相對狹窄,如B19只能在極少的幾個細胞系中復制,且效力很低。目前還未有人博卡病毒感染性克隆及其敏感細胞系的報道,成為對該病毒感染機制研究的瓶頸。本實驗從呼吸道疾病患兒痰液中鑒定出人博卡病毒,并構建了病毒基因組克隆 (缺少 ITR),基因序列分析表明為HBoV1型。構建了病毒啟動子活性檢測重組載體,轉染多種哺乳動物細胞,實驗證明HBoV1啟動子在所試哺乳動物細胞中具有很高活性,為進一步研究 HBoV1的轉錄、翻譯等分子機制提供重要平臺。
1 材料與方法
1.1 菌株、質粒及細胞
大腸桿菌菌株DH10B、質粒pBluescript SKⅡ、pGL3-Basic、pEGFP-N1由本實驗室保存;人胚腎上皮細胞293T/293GP2P6、人肺成纖維細胞WI-38AV、人肺癌細胞A-549、兔腎細胞RK13、豬鼻甲骨黏膜成纖維細胞PT、HeLa細胞由本實驗室保存。
1.2 主要試劑
限制性內切酶 (EcoRⅠ、SalⅠ、XbaⅠ、HindⅢ、BglⅡ、NcoⅠ)、Taq DNA 聚合酶、T4 DNA聚合酶、DNA 連接酶均購自 TaKaRa公司;DNA片段膠回收試劑盒購自博大泰克公司;DNA marker、高純度質粒小提中量試劑盒購自天根 (Tiangen) 生物技術公司;TA Cloning試劑盒、轉染試劑Lipofectamine Reagent購自Invitrogen公司;熒光素酶檢測試劑盒Luciferase Assay System購自Promega公司;QIAmp DNA Mini Kit購自Qiagen公司。DMEM購自Gibico公司;胎牛血清購自Hyclone公司。
1.3 樣品處理及病毒DNA提取
本實驗 941例痰液樣品均采自湖北省婦幼保健院具有呼吸道感染癥狀的患兒。在痰樣中加入 5倍體積的生理鹽水,去除唾液,然后加入等體積化痰劑 (含0.1% DTT) 混勻,37 ℃水浴30 min后1 500 r/min離心10 min,取上清。取200 μL提取病毒DNA,剩余上清?70 ℃保存。QIAmp DNA Mini Kit提取病毒基因組DNA,操作按照說明書進行。
1.4 PCR檢測
根據 GenBank上已公布的人博卡病毒基因組序列 DQ000496[1]設計引物 (Forward: 5′-TATCGT CTTGCACTGCTTCG-3′; Reverse: 5′-AGAGTAGGC GTGATCATGTAA-3′),以提取的痰樣病毒DNA為模板,擴增保守區 NP1基因,引物合成由北京奧科生物公司完成。反應條件:94 ℃預變性3 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min ,共35個循環;72 ℃延伸10 min。PCR產物經0.8%瓊脂糖凝膠電泳檢測。
1.5 HBoV1基因組克隆的構建
取一管已鑒定為HBoV陽性的痰樣DNA為模板,根據 DQ000496基因組測序結果,結合pBluescript SKⅡ載體的酶切位點,對基因組分 3段擴增:a段 (1 408 bp)、b段 (2 893bp) 和c段(997 bp),引物見表1。利用瓊脂糖凝膠DNA回收試劑盒回收3段PCR產物。先將a和c段與T載體連接,轉化DH10B感受態細胞,挑取克隆并鑒定,b段PCR擴增產物通過HindⅢ和EcoRⅠ酶切位點插入pBluescript SKⅡ載體,再將a和c段分別通過SalⅠ/HindⅢ、EcoRⅠ/XbaⅠ酶切位點插入上述質粒中,構建HBoV1基因組克隆,酶切鑒定重組質粒。
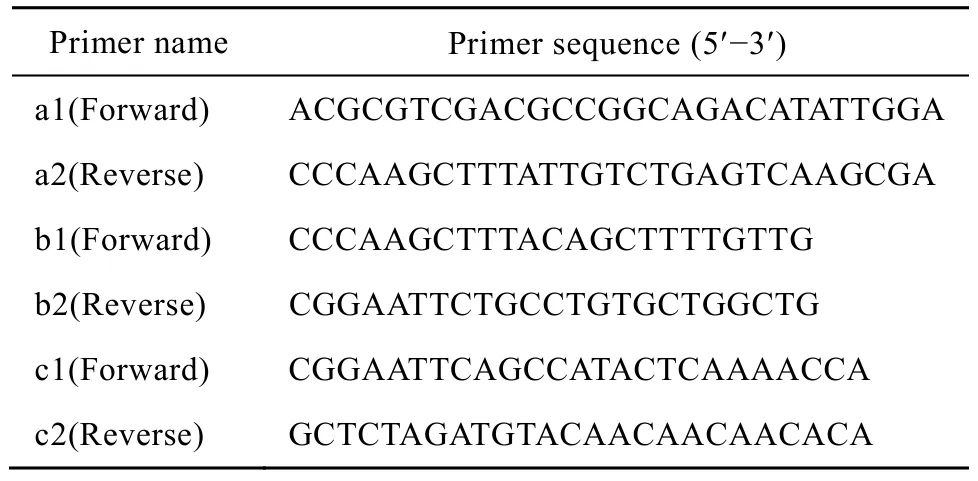
表1 構建HBoV1基因組克隆所用引物Table 1 Primers used for construction of a full-length genome clone of HBoV1
1.6 HBoV1基因組測序及序列分析
將已連T載體的a、c段及插入pBluescript SKⅡ載體的 b段分別測序,測序工作由北京奧科生物公司完成,采用 Phrap拼接基因組測序片段。采用Sequin完成序列注釋和測序提交文件的生成,測序后拼裝的基因組命名為WHL-1。利用DNAStar軟件對已測 HBoV1基因組序列和推導的氨基酸序列進行分析。
1.7 HBoV1啟動子重組載體的構建
根據 GU139423序列設計啟動子上下游引物,分別引入 BglⅡ、HindⅢ酶切位點:上游引物:5′-GAAGATCTGCCGGCAGACATATTC-3′,下游引物:5′-CCCAAGCTTAGTAGACTCACCACAAGCG T-3′。以 WHL-1質粒為模板進行 PCR擴增,反應條件:94 ℃預變性3 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min ,共30個循環;72 ℃延伸10 min。PCR產物經0.8%瓊脂糖凝膠電泳檢測。利用瓊脂糖凝膠DNA回收試劑盒回收 PCR產物。重組載體pGL3-Basic-pBoV的構建:分別用限制性內切酶BglⅡ和HindⅢ對回收片段和pGL3-Basic載體質粒進行雙酶切,酶切回收產物16 ℃連接過夜,轉化大腸桿菌DH10B感受態細胞,挑取陽性克隆搖菌,酶切鑒定后測序。重組載體pGL3-pBoV-EGFP的構建:以pEGFP-N1質粒為模板擴增EGFP基因片段,通過酶切連接將EGFP基因替換pGL3-Basic-pBoV載體上的熒光素酶基因 (luc+),酶切鑒定后測序。陽性對照重組載體 pGL3-Basic-pCMV的構建:以pEGFP-N1質粒為模板擴增 CMV啟動子區,插入pGL3-Basic載體上的BglⅡ和HindⅢ酶切位點之間,酶切鑒定后測序。
1.8 HBoV1啟動子活性檢測
1.8.1 熒光顯微鏡檢測GFP的表達
將試驗細胞 (293T/293GP2P6、WI-38AV、A-549、RK13、PT、HeLa) 接種在24孔板中,37 ℃、5% CO2培養箱中過夜培養。在細胞長至 80%時,用PBS洗2遍細胞,加入無血清培養基,同時按1∶2 (μg∶μL)的比例將質粒pGL3-pBoV-EGFP與轉染試劑Lipofectamine混合 (每孔加入1 μg質粒和2 μL轉染試劑)。室溫放置30 min后將混合物加入清洗過的細胞中,在37 ℃、5% CO2培養箱中培養4 h,用含 10%小牛血清的 DMEM 更換培養基繼續培養,以pEGFP-N1質粒轉染各種細胞為陽性對照。24 h后熒光顯微鏡觀察。
1.8.2 熒光素酶報告基因活性測定
將試驗細胞 (293T、WI-38AV、A-549、HeLa)按 1×104個細胞/孔的量接種在 96孔板中,質粒pGL3-Basic-pBoV轉染細胞按照上述操作方法進行。以 pGL3-Basic-pCMV質粒轉染各種細胞為陽性對照。培養24 h后吸棄細胞培養基,用PBS洗1次,加入裂解液20 μL/孔,室溫搖動15 min。按照熒光素酶檢測試劑盒 (Luciferase Assay System) 的操作說明書,每孔加入100 μL LAR反應液,混勻后加入96孔檢測板,設定微孔板式發光儀 (Centro LB 960)延遲2 s,發光儀測讀10 s,測讀螢火蟲熒光素酶發光值。轉染試驗重復3次,數據采用Origin6.0軟件進行統計學分析。
2 結果與分析
2.1 HBoV1陽性樣本的PCR檢測
PCR擴增產物表現為單一特異條帶,分子量約為600 bp,與預期結果相符 (圖1)。PCR產物回收后連T載體測序,利用NCBI網站的Blast工具對測序結果進行比對分析表明為HBoV NP1基因,該基因在博卡病毒屬中非常保守,成為博卡病毒鑒定常用目標基因。941例痰液樣本檢測到33例HBoV陽性擴增產物,陽性率為3.51% (33/941)。33例陽性患者臨床診斷為肺炎或支氣管肺炎,且大多為 1歲以下患兒 (24例),占整個患者陽性樣的72.7%;為人類博卡病毒的易感人群。
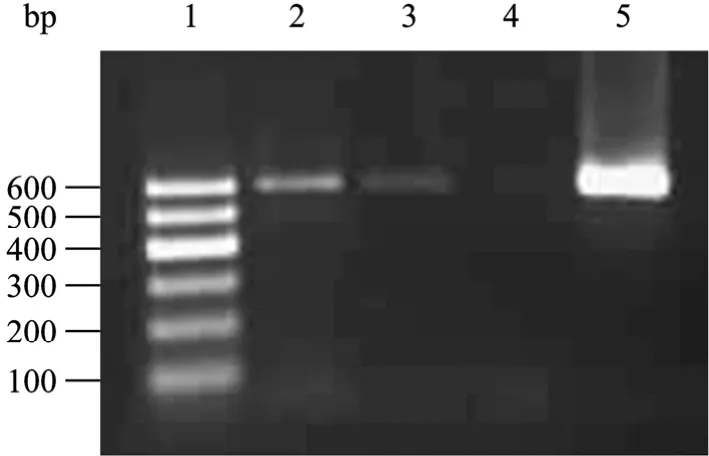
圖1 HBoV1 NP基因的PCR產物鑒定Fig. 1 PCR products of HBoV1. 1: DNA marker I; 2,3,5: positive samples; 4: negative sample.
2.2 HBoV1基因組克隆的構建
以HBoV陽性樣本DNA為模板,利用PCR方法分 3段擴增病毒基因組,通過瓊脂糖凝膠電泳得到與預期擴增片段大小一致的3條產物:a段(1 408 bp)、b段 (2 893 bp) 和c段 (997 bp) (圖2A)。按照方法 1.5構建病毒基因組克隆,雙酶切鑒定如圖2B所示:SalⅠ+XbaⅠ酶切下基因全長 (a+b+c);SalⅠ+HindⅢ酶切下a段;HindⅢ+EcoRⅠ酶切下b段;EcoRⅠ+XbaⅠ酶切下c段。初步驗證成功構建了人博卡病毒基因組,命名為WHL-1。
2.3 HBoV1基因組測序及序列分析
對已鑒定的含有病毒基因組各個片段的質粒進行測序,測序結果拼接后最終獲得HBoV基因組中間大片段序列,該序列全長5 299 bp,編碼3個開放閱讀框:NS1、NP1和VP1/VP2,GenBank登錄號為GU139423,基因型為HBoV1型。該序列與已報道序列DQ000496氨基酸同源性達到99%,僅16個核苷酸發生變異,大多位于易突變的衣殼蛋白編碼區,而NP1基因完全保守,其他細小病毒中并未發現與NP1同源的蛋白。NP1在MVC病毒DNA復制過程中發揮重要作用,缺失NP1基因的MVC可由HBoV1或 BPV1的 NP1基因回補,正常進行病毒DNA復制[15]。對病毒基因組啟動子區分析顯示,該啟動子中心區域位于146~196 nt,距離NS1起始密碼子僅91個核苷酸。該片段包含真核生物基因啟動子的基本元件TATA盒、CAAT框以及轉錄起始基序 (圖3),這些元件是否具備功能還需進一步驗證。
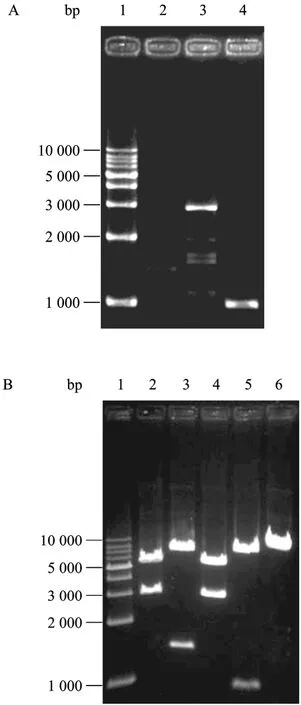
圖2 HBoV1基因組PCR分段擴增 (A) 及重組載體的酶切鑒定 (B)Fig. 2 PCR products of three segments (A) and recombinant plasmid identified by enzyme digestion (B). (A) 1: 1 kb DNA marker; 2?4: PCR product of fragments from a to c. (B) 1: 1 kb DNA marker; 2: pSK-a-b-c digested with Sal I and Xba I; 3: pSK-a-b-c digested with Sal I and Hind III; 4: pSK-a-b-c digested with Hind III and EcoR I; 5: pSK-a-b-c digested with EcoR I and Xba I; 6: pSK-a-b-c digested with EcoR I.

圖3 HBoV1啟動子序列Fig. 3 Nucleotide sequence of the HBoV1 promoter.
2.4 HBoV1啟動子重組載體的構建
以WHL-1質粒為模板,PCR擴增HBoV1啟動子序列 (1~252 nt),凝膠電泳檢測大小正確(圖4A2),PCR回收片段與pGL3-Basic載體連接,酶切鑒定,切下250 bp大小條帶 (圖4A3),與預期結果一致,成功構建了pGL3-Basic-pBoV重組載體。以pEGFP-N1質粒為模板擴增EGFP基因片段,大小為719 bp (圖4B2),通過雙酶切用EGFP基因替換已構建載體pGL3-Basic-pBoV的luc報告基因,酶切鑒定如圖3B3。以pEGFP-N1質粒為模板擴增CMV啟動子區,大小589 bp (圖4C2),成功構建陽性對照重組載體pGL3-Basic-pCMV,酶切鑒定如圖4C3。構建載體的測序工作由南京金斯瑞生物有限公司完成。
2.5 HBoV1啟動子活性檢測
重組質粒pGL3-pBoV-EGFP轉染試驗細胞24 h后,熒光顯微鏡觀察細胞中 EGFP蛋白的表達 (圖5),以pEGFP-N1載體轉染各試驗細胞為陽性對照。所有試驗細胞均檢測到綠色熒光,表明 HBoV1啟動子在哺乳動物細胞中啟動下游報告基因EGFP的轉錄、表達。pEGFP-N1是常用的帶有EGFP標簽的表達載體,在CMV的啟動下,外源基因與EGFP融合表達,在熒光顯微鏡下可直接觀察蛋白表達。各細胞試驗組與對照細胞組相比,熒光強度相當甚至更強,顯示 HBoV1啟動子在哺乳動物中具有很強活性。293T、293G和PT細胞試驗組中表達熒光蛋白的細胞明顯多于 WI-38AV、A-549、RK13和HeLa細胞試驗組,原因在于Lipofectamine Reagent介導下對各種細胞轉染效率不同。同時,我們采用熒光素酶檢測系統對啟動子活性定量檢測,以pGL3-Basic-pCMV轉染各種試驗細胞測得發光值為1,HBoV1啟動子在4種細胞中熒光素酶活性相對值見圖 6,說明啟動子在不同細胞中活性都比CMV啟動子強 (熒光素酶相對活性都大于 1)。在293T、HeLa、A-549和WI-38AV細胞中,HBoV1啟動子活性分別是CMV的4.5倍、2.2倍、1.4倍和1.1倍。
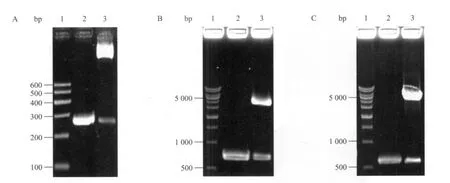
圖4 啟動子重組載體的構建Fig. 4 Construction of HBoV1 promoter vectors. (A) 1: DNA marker I; 2: PCR product of HBoV1 promoter; 3: pGL3-Basic-pBoV digested with Bgl II/Hind III. (B) 1: 1 kb DNA marker; 2: PCR product of EGFP; 3: pGL3-pBoV-EGFP digested with Nco I/Xba I. (C) 1: 1 kb DNA marker; 2: PCR product of CMV promoter. 3: pGL3-Basic-pCMV digested with Bgl II/Hind III.
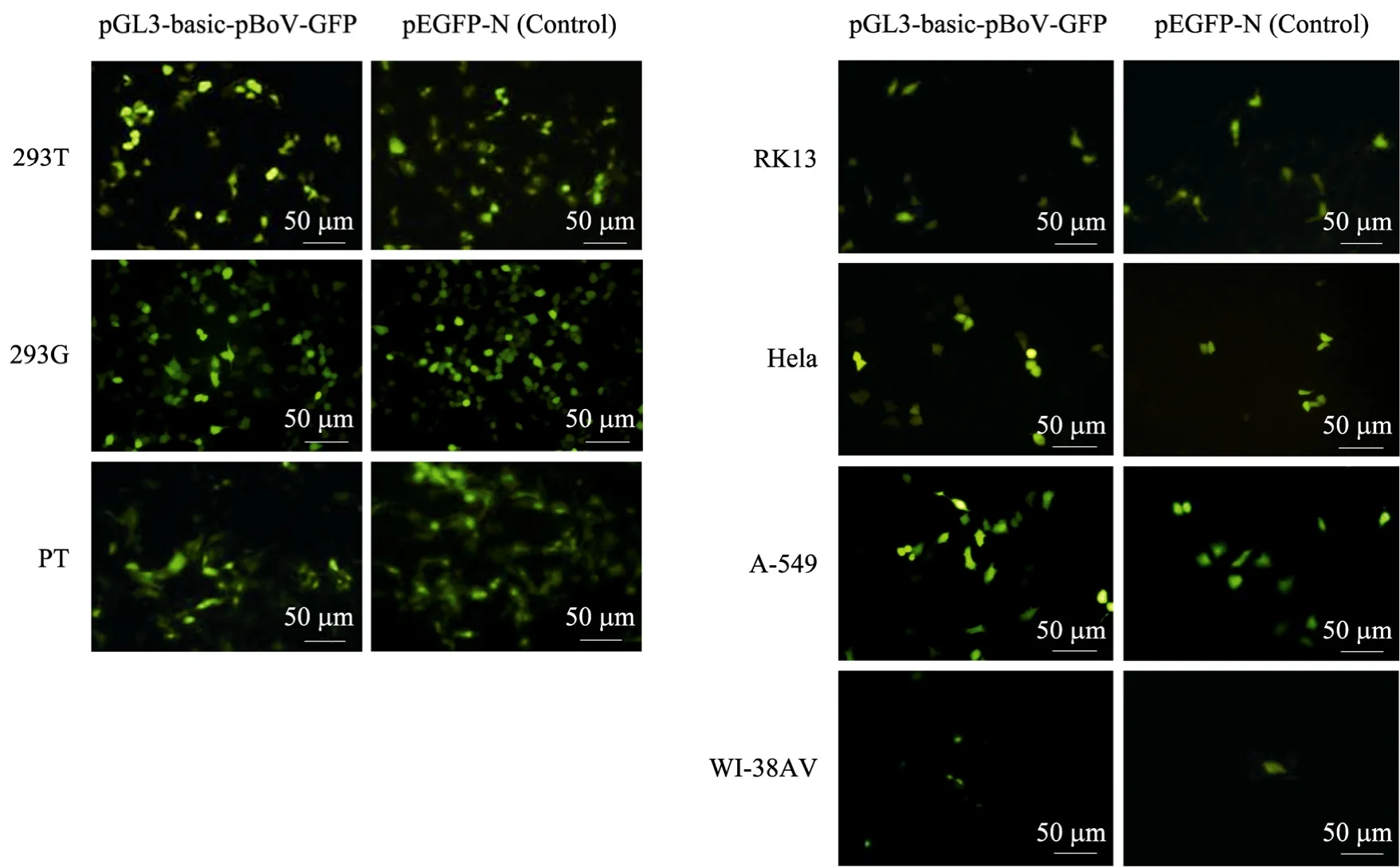
圖5 熒光顯微鏡檢測HBoV1啟動子下的EGFP在不同哺乳動物細胞中的表達Fig. 5 Detection the EGFP expression by HBoV1 promoter function in different cell lines with fluorescence microscope (bar=50 μm).
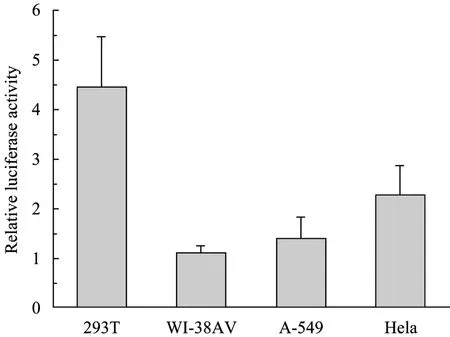
圖6 HBoV1啟動子在4種細胞中的活性比較Fig. 6 Comparison of the strengths of HBoV1 promoter in different cell lines. The values were calculated by dividing the amount of relative luciferase activity driven by HBoV1 promoter. The activity of the CMV promoter was set at 1 in all cell types. The error bars indicate standard deviations of 3 experiments and each performed in triplicate.
3 討論
人博卡病毒是近年來新發現的感染人類的細小病毒,最初是在患有呼吸道疾病的小兒鼻咽抽吸物中篩選出該病毒,且在世界范圍內都有發現該病毒感染。近兩年又在患有胃腸道疾病患者排泄物中鑒定出不同基因型的 HBoV。在成人的呼吸道分泌物中偶爾也能檢測到人博卡病毒[16]。本研究采集的標本來自患有呼吸道疾病的患兒,941例痰液樣本檢測到33例HBoV陽性擴增產物,其中1歲以下嬰幼兒患者占陽性樣72.7%,為人博卡病毒的易感人群,而有報道5歲兒童90%以上都具有人博卡病毒抗體[17]。
本實驗構建了人博卡病毒基因組克隆,對其測序并與已報道的序列比對發現,該博卡病毒屬于HBoV1型。與其他細小病毒類似,博卡病毒基因組兩端具有ITR結構,但迄今為止還未能得到該序列,也未見有相關報道。我們已構建的病毒重組載體WHL-1也只包括中間5 299 nt的大片段,由于缺乏ITR結構,采用Southern blotting方法,在轉染WHL-1的293T細胞中并未檢測到病毒DNA復制 (結果未顯示),進一步證明ITR結構對病毒復制是必需的。由于未分離得到HBoV病毒粒子,感染性克隆的構建又受阻于ITR結構,對人類博卡病毒的研究多局限于流行病學研究。獲得感染性克隆是研究病毒感染的分子機制的突破口,我們正進行這方面的工作。
人類博卡病毒唯一啟動子位于基因組左端,由于缺乏ITR結構,啟動子區是否會受到影響而無活性,能否在哺乳動物細胞中行使功能。我們將基因組左端252個核苷酸克隆到pGL3-Basic上,分別轉染人類、豬、兔代表細胞系,結果顯示,在這幾類哺乳動物細胞中,HBoV1啟動子都具有活性,且在293T細胞中活性最強。首次證明HBoV1啟動子在大多數哺乳動物細胞中具有較高活性,為深入研究博卡病毒轉錄和翻譯機制提供了很好的平臺。
[1] Allander T, Tammi MT, Eriksson M, et al. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proc Natl Acad Sci USA, 2005, 102(36):12891?12896.
[2] Vicente D, Cilla G, Montes M, et al. Human bocavirus, a respiratory and enteric virus. Emerg Infect Dis, 2007, 13(4): 636?637.
[3] Allander T, Jartti T, Gupta S, et al. Human bocavirus and acute wheezing in children. Clin Infect Dis, 2007, 44(7): 904?910.
[4] Kesebir D, Vazquez M, Weibel C, et al. Human bocavirus infection in young children in the United States: molecular epidemiological profle and clinical characteristics of a newly emerging respiratory virus. J Infect Dis, 2006, 194(9):1276?1282.
[5] Manning A, Russell V, Eastick K, et al. Epidemiological profle and clinical associations of human bocavirus and other human parvoviruses. J Infect Dis, 2006, 194(9): 1283?1290.
[6] Kapoor A, Slikas E, Simmonds P, et al. A newly identifed bocavirus species in human stool. J Infect Dis, 2009, 199(2): 196?200.
[7] Arthur JL, Higgins GD, Davidson GP, et al. A novel bocavirus associated with acute gastroenteritis in Australian children. PLoS Pathog, 2009, 5(4): e1000391.
[8] Kapoor A, Simmonds P, Slikas E, et al. Human bocaviruses are highly diverse, dispersed, recombination prone, and prevalent in enteric infections. J Infect Dis, 2010, 201(11): 1633?1643.
[9] Dijkman R, Koekkoek SM, Molenkamp R, et al. Human bocavirus can be cultured in differentiated human airway epithelial cells. J Virol, 2009, 83(15): 7739?7748.
[10] Ozawa K, Ayub J, Hao YS, et al. Novel transcription map for the B19 (human) pathogenic parvovirus. J Virol, 1987, 61(8): 2395?2406.
[11] Liu ZW, Qiu JM, Cheng F, et al. Comparison of the transcription profile of simian parvovirus with that of the human erythrovirus B19 reveals a number of unique features. J Virol, 2004, 78(23): 12929?12939.
[12] Qiu JM, Cheng F, Burger LR, et al. The transcription profile of aleutian mink disease virus (AMDV) in CRFK cells is generated by alternative processing of pre-mRNAs produced from a single promoter. J Virol, 2006, 80(2): 654?662.
[13] Burnett E, Cotmore SF, Tattersall P, et al. Segregation of a single outboard left-end origin is essential for the viability of parvovirus minute virus of mice. J Virol, 2006, 80(21): 10879?10883.
[14] Chen AY, Cheng F, Lou S, et al. Characterization of the gene expression profile of human bocavirus. Virology, 2010, 403(2):145?154.
[15] Sun YN, Chen AY, Cheng F, et al. Molecular characterization of infectious clones of the minute virus of canines reveals unique features of bocaviruses. J Virol, 2009, 83(8): 3956?3967.
[16] Longtin J, Bastien M, Gilca R, et al. Human bocavirus infections in hospitalized children and adults. Emerg Infect Dis, 2008, 14(2): 217?221.
[17] Endo R, Ishiguro N, Kikuta H, et al. Seroepidemiology of human bocavirus in Hokkaido prefecture, Japan J Clin Microbiol, 2007, 45(10): 3218?3232.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
