三疣梭子蟹Profilin基因全長cDNA的克隆與表達分析
2010-12-25 06:40:08王日昕
Zoological Research 2010年3期
關(guān)鍵詞:分析
申 望, 葉 茂, 石 戈, 王日昕
(浙江海洋學(xué)院 海洋科學(xué)學(xué)院海洋生物資源及分子工程實驗室,浙江 舟山 316004)
三疣梭子蟹Profilin基因全長cDNA的克隆與表達分析
申 望, 葉 茂, 石 戈, 王日昕*
(浙江海洋學(xué)院 海洋科學(xué)學(xué)院海洋生物資源及分子工程實驗室,浙江 舟山 316004)
三疣梭子蟹(Portunus trituberculatus)是我國沿海重要養(yǎng)殖品種之一。近年來,養(yǎng)殖病害呈逐年上升趨勢,制約了三疣梭子蟹養(yǎng)殖產(chǎn)業(yè)的健康可持續(xù)發(fā)展。克隆三疣梭子蟹免疫相關(guān)基因、研究免疫基因的功能和作用機制,可為三疣梭子蟹養(yǎng)殖病害的防治奠定基礎(chǔ)。本研究從三疣梭子蟹血細胞全長cDNA文庫中克隆了742 bp的profilin基因全長cDNA。該profilin全長cDNA中開放閱讀框長375 bp,編碼125 aa。推導(dǎo)的三疣梭子蟹profilin理論等電點pI 5.87,氨基酸序列與岡比亞按蚊(Anopheles gambiae)profilin同源性最高,序列一致性為42.9%。熒光定量RT-PCR分析結(jié)果顯示,在正常的三疣梭子蟹機體中,血細胞profilin表達水平最高,其次為肝胰臟;在致病菌副溶血弧菌(Vibrio parahaemolyticus)誘導(dǎo)后,血細胞中profilin表達量顯著上升(P<0.01),表明profilin可能參與了三疣梭子蟹的免疫防御反應(yīng),是一個免疫相關(guān)因子。
三疣梭子蟹;免疫;基因克隆;Profilin;基因表達
Profilin最初是從牛脾中純化鑒定的一個與肌動蛋白單體(G-actin)結(jié)合的相對分子質(zhì)量15 k的小分子蛋白質(zhì)。現(xiàn)有研究表明,profilin普遍存在于真核生物細胞中(Carlsson et al, 1977; Jockusch et al, 2007),與肌動蛋白單體按1:1結(jié)合,催化肌動蛋白單體的ADP/ATP核苷交換,生成G-ATP-actin,調(diào)節(jié)肌動蛋白的動力學(xué)活性,影響運動細胞邊緣肌動蛋白網(wǎng)絡(luò)的組裝、維持和去組裝(Haugwitz et al, 1994; Pollard & Borisy, 2003; Romero et al, 2007),參與肌動蛋白活性基礎(chǔ)上的生命活動,如細胞運動、黏附、分裂、形態(tài)發(fā)生以及細胞內(nèi)的膜泡運輸?shù)取rofilin除參與肌動蛋白相關(guān)生理活動外,還參與機體的免疫防御反應(yīng),是一個免疫相關(guān)因子,如海膽體腔細胞經(jīng)脂多糖誘導(dǎo)后,profilin轉(zhuǎn)錄水平顯著提高(Smith et al, 1995);人乳腺癌細胞中profilin表達量顯著低于正常乳腺上皮細胞中profilin的表達量,通過轉(zhuǎn)染profilin cDNA提高乳腺癌細胞中profilin表達量后,乳腺癌細胞恢復(fù)正常生長(Janke et al, 2000);果蠅profilin基因突變的血細胞吞噬活性增強,表明果蠅profilin是一個關(guān)鍵的吞噬活性調(diào)節(jié)因子(Pearson et al, 2003)。
三疣梭子蟹(Portunus trituberculatus)是我國重要的海產(chǎn)經(jīng)濟蟹類,也是90年代蝦病暴發(fā)和流行后,作為對蝦替代養(yǎng)殖品種發(fā)展起來的大宗養(yǎng)殖品種。但近年來由于養(yǎng)殖規(guī)模的不斷擴大以及養(yǎng)殖集約化程度的不斷提高,三疣梭子蟹養(yǎng)殖病害也頻頻爆發(fā),制約了三疣梭子蟹養(yǎng)殖業(yè)的進一步發(fā)展(Chen et al, 2005; Li et al, 2008)。克隆三疣梭子蟹免疫相關(guān)基因,研究免疫基因的功能和作用機制,可為三疣梭子蟹養(yǎng)殖病害的防治以及遺傳育種奠定良好基礎(chǔ)。本研究從三疣梭子蟹血細胞全長cDNA文庫中克隆了一個profilin基因的全長cDNA序列,對其進行生物信息學(xué)分析,并應(yīng)用熒光定量RT-PCR技術(shù)研究了該基因在正常三疣梭子蟹不同組織中的表達水平差異以及致病菌誘導(dǎo)前后血細胞中的表達水平變化,為研究profilin參與的三疣梭子蟹免疫防御反應(yīng)機制奠定基礎(chǔ)。
1 材料與方法
1.1 三疣梭子蟹血細胞全長cDNA文庫的構(gòu)建及profilin基因克隆
三疣梭子蟹采自浙江舟山海域。成年雄性三疣梭子蟹采集后飼養(yǎng)于恒溫水族箱(25℃),飼養(yǎng)24 h后,以內(nèi)含抗凝劑(0.45 mol/L NaCl,0.1 mol/L葡萄糖,26 mmol/L檸檬酸,30 mmol/L檸檬酸鈉,10 mmol/L EDTA,pH4.6)的注射器從蟹肢體未硬化部位抽取血淋巴,立即4℃ 2 000 g離心5 min,收集血細胞(S?derh?ll & Smith, 1983),采用TRIZOL法提取三疣梭子蟹血細胞總RNA,mRNA的分離根據(jù)Invitrogen公司FastTrack? 2.0 Kit試劑盒操作手冊進行,通過poly(T)纖維素柱親和層析,從三疣梭子蟹血細胞總RNA中分離poly A+mRNA。全長cDNA文庫的構(gòu)建按Clontech公司SMART cDNA全長文庫構(gòu)建試劑盒說明書進行,連接載體使用pGEM-T easy T載體,轉(zhuǎn)化菌株為E. coli(DH10B)。文庫克隆子測序引物為M13F (5'-TGT AAA ACG ACG GCC AGT-3')和M13R(5'-CAG GAA ACA GCT ATG ACC-3')。測得的ESTs序列通過BLASTX檢索GenBank nr數(shù)據(jù)庫尋找同源基因。
1.2 三疣梭子蟹profilin基因的生物信息學(xué)分析與系統(tǒng)樹構(gòu)建
在線計算三疣梭子蟹profilin理論分子量及等電點(http://us.expasy.org/ tools/ pi_tool.html; Bjellqvist et al, 1993; Bjellqvist et al, 1994);NetPhos2.0 Server在線分析磷酸化位點(http://www.cbs.dtu.dk/services/NetPhos/; Blom et al, 1999);TargetP 1.1 Server在線分析亞細胞定位信息(http://www.cbs.dtu.dk/services/TargetP/; Emanuelsson et al, 2000);氨基酸序列比對使用Clustal W(Thompson et al, 1994);以MEGA 4.1軟件包(Kumar et al, 2008)中的鄰接法(nerghbor-joining method, NJ)構(gòu)建節(jié)肢動物profilin系統(tǒng)樹,選擇人profilin為外群,參數(shù)設(shè)置為默認的泊松校正算法,不考慮插入/缺失位點,同時,應(yīng)用自舉檢驗(bootstrap test)重抽樣1 000次估計系統(tǒng)樹中節(jié)點的自引導(dǎo)值(bootstrap value)。
構(gòu)建系統(tǒng)樹所用節(jié)肢動物和人profilin如下:中國明對蝦(Fenneropenaeus chinensis,F(xiàn)J480175)、凡納濱對蝦(Litopenaeus vannamei,ABI93174)、斑節(jié)對蝦(Penaeus monodon,ABU97474)、家蠶(Bombyx mori,NP_001037108)、黑腹果蠅(Drosophila melanogaster,NP_477016)、岡比亞按蚊(Anopheles gambiae,XP_553744)、蚜蟲(Acyrthosiphon pisum,NP_001156129、XP_ 001945869、XP_001945923)、魚虱(Lepeophtheirus salmonis,ACO12777)、魚虱(Caligus rogercresseyi,ACO10829)、黑腳硬蜱(Ixodes scapularis,XP_002414739)、無爪螨(Blomia tropicalis,AAQ24553)、棉蘭皺皮螨(Suidasia medanensis,AAX34044)、人(Homo sapiens,NP_005013、NP_444252、NP_002619、NP_001025057、NP_955378)。
1.3 Profilin基因表達的熒光定量RT-PCR分析
1.3.1 Profilin基因在三疣梭子蟹不同組織中的表達分析 從5只雄性三疣梭子蟹分別采集等量的血細胞、肌肉、肝胰臟、眼組織,按組織差別將5個體來源的組織合并后提取總RNA。血細胞總RNA提取,按1.1進行;其它組織采集后先液氮研磨,再用TRIZOL 法提取總RNA。以組織總RNA為模板,oligo(dT)18為引物, 按全式金EasyScript Reverse Transcriptase說明書操作反轉(zhuǎn)錄合成cDNA第一鏈。以三疣梭子蟹核糖體蛋白S18(RpS18)為內(nèi)參基因,采用熒光定量RT-PCR技術(shù)研究profilin基因在不同組織中的差異表達。 RpS18基因RT-PCR引物:Ptrs18-F:5'-AGG AGG AGG TTG AGA AGA TTG T-3',Ptrs18-R:5'- GCA GCT TGG TTT CCA GGT AG-3', 擴增片段長度141 bp;profilin基因RT-PCR引物:Ptpro-F:5'-GCA AGA AGG AAA CAA CTG GAG -3',Ptpro-R:5'-GAG GTG GTC AGC CAT ACA GTC-3',擴增片段長度125 bp。RT-PCR反應(yīng)體系: 第1鏈cDNA 2μL,SYBR Premix Ex TaqTM(2 ×)12.5μL,上游引物0.5 μL(10 μmol/L),下游引物0.5 μL (10 μmol/L),加9.5 μL去離子水至總體積25 μL。擴增條件:95℃預(yù)變性2 min;94℃變性10 s,60℃ 40 s,40個循環(huán)。反應(yīng)在ABI 7500Fast實時定量PCR儀上進行,定量的數(shù)據(jù)結(jié)果采用2-△△Ct法(Livak et al, 2001)進行分析。三疣梭子蟹profilin在不同組織及致病菌誘導(dǎo)前后的相對表達量采用SPSS統(tǒng)計軟件進行Ttest檢驗,P<0. 05認為差異有統(tǒng)計學(xué)意義。
1.3.2 三疣梭子蟹血細胞Profilin基因在副溶血弧菌(Vibrio parahaemolyticus)誘導(dǎo)下表達的時空差異分析 副溶血弧菌在LB液體培養(yǎng)基擴大培養(yǎng)后離心收集細菌,無菌去離子水洗滌3次,最后用無菌去離子水稀釋到109CFU/mL備用。5只雄性三疣梭子蟹(重250~300 g/只)在恒溫水族箱(25℃)暫養(yǎng)24 h后,每只抽取0.3 mL血淋巴,合并后按1.1提取總RNA,然后每只蟹注射50 μL菌液,并在注射菌液后3、6、12、24 h如前所述抽取血淋巴并提取總RNA。實時熒光定量PCR分析操作按1.3.1進行。
2 結(jié) 果
2.1 三疣梭子蟹profilin基因克隆與鑒定
用引物M13F和M13R對三疣梭子蟹全長cDNA文庫進行初步測序,獲得74條ESTs序列,將ESTs序列進行拼接,共得到23條unigene,包含6個序列重疊群(Contigs)和17條Singletons。其中一個包含7條ESTs序列的序列重疊群經(jīng)BLASTX檢索到的同源基因均為profilin,并與岡比亞按蚊profilin(XP_553744)同源性最高,氨基酸序列一致性為42.9%,因此,該基因為三疣梭子蟹的profilin基因。三疣梭子蟹血細胞cDNA文庫測序得到的7條profilin ESTs序列僅在3'-UTR末端可能由于加尾時剪切位點不同形成1~3個核苷酸的長度差異,因此,7條profilin ESTs序列應(yīng)為同一基因的轉(zhuǎn)錄產(chǎn)物。三疣梭子蟹profilin基因cDNA一致序列長度為742 bp(不包括polyA,Genbank登錄號:GU253912),由66 bp的5'-UTR、375 bp的編碼區(qū)和301 bp的3'-UTR組成,在3'-UTR末端有加尾信號AATAAA(圖1)。編碼區(qū)推導(dǎo)的氨基酸序列由125個氨基酸殘基組成,理論相對分子質(zhì)量13 739.38,預(yù)期等電點pI 5.87,在線磷酸化位點分析顯示三疣梭子蟹profilin共有11個可能的磷酸化位點,分別為Ser5/32/38/56/58/71/104/123、Thr84和Tyr6/125。

圖 1 三疣梭子蟹profilin的全長cDNA序列和推導(dǎo)的氨基酸序列Fig. 1 The full-length cDNA sequence and deduced amino acid sequence of the profilin from Portunus trituberculatus
蛋白亞細胞定位分析結(jié)果表明,推導(dǎo)的三疣梭子蟹profilin蛋白無N 末端信號肽、無線粒體及核定位信號、無內(nèi)質(zhì)網(wǎng)停留信號、無過氧化酶體結(jié)合信號、無DNA或RNA結(jié)合信號、無外分泌信號,表明該profilin蛋白分布于細胞質(zhì)中,細胞定位特征與引言中所述功能一致。
2.2 Profilin的序列分析比對與系統(tǒng)發(fā)生分析
推導(dǎo)的三疣梭子蟹profilin氨基酸序列與6種節(jié)肢動物門動物profilin的序列比對結(jié)果如圖2所示(圖2)。三疣梭子蟹profilin與中國明對蝦(Fenneropenaeus chinensis,F(xiàn)J480175)、凡納濱對蝦(Litopenaeus vannamei,ABI93174)、斑節(jié)對蝦(Penaeus monodon,ABU97474)、家蠶(Bombyx mori,NP_001037108)、黑腹果蠅(Drosophila melanogaster,NP_477016)和岡比亞按蚊(Anopheles gambiae,XP_553744)等profilin的氨基酸一致性分別為35.71 %、36.5 %、34.9 %、40.5 %、38.10 %和42.9 %。
以人的5種profilin亞型為外群構(gòu)建的節(jié)肢動物門profilin NJ系統(tǒng)樹如圖3所示。有螯亞門(Chelicerata)3個物種的profilin聚在一枝;有顎亞門(Mandibulata)昆蟲綱代表物種的profilin聚在一枝,甲殼綱文獻報道的5種profilin以及本研究報道的三疣梭子蟹profilin也聚為單獨一枝;但在甲殼綱中,3種對蝦profilin并未聚為一枝,其中凡納濱對蝦(軟甲亞綱,Malacostraca)profilin與顎足亞綱(Maxillopoda)兩種魚虱的profilin親緣關(guān)系更近;三疣梭子蟹與對蝦同屬軟甲亞綱十足目(Decapoda),三疣梭子蟹profilin與中國明對蝦、斑節(jié)對蝦profilin親緣關(guān)系較近,而與凡納濱對蝦profilin親緣關(guān)系較遠。
2.3 三疣梭子蟹profilin的表達分析
熒光定量RT-PCR分析結(jié)果顯示,profilin在未誘導(dǎo)的三疣梭子蟹血細胞、肝胰臟、眼和肌肉中均有表達(圖4A)。其中血細胞profilin表達量最高,其次為肝胰臟,相對表達量為血細胞的0.2倍(P<0.01),而在眼和肌肉中profilin表達量極低,相對表達量分別僅為血細胞的0.028倍(P<0.01)和0.002倍(P<0.01)。
以誘導(dǎo)前三疣梭子蟹血細胞profilin表達量為對照,熒光定量RT-PCR分析副溶血弧菌誘導(dǎo)后不同時間血細胞profilin表達量變化結(jié)果顯示:誘導(dǎo)3、6、12和24 h后,profilin表達量分別相當(dāng)于與誘導(dǎo)前的1.91倍(P<0.01)、1.21倍(P<0.01)、0.78倍(P<0.01)和0.62倍(P<0.01),表明血細胞中profilin的在副溶血弧菌誘導(dǎo)下表達量上調(diào),后下調(diào)(圖4B)。

圖 2 節(jié)肢動物代表性profilin氨基酸序列比對Fig. 2 Multiple sequence alignment of the profilins from arthropod
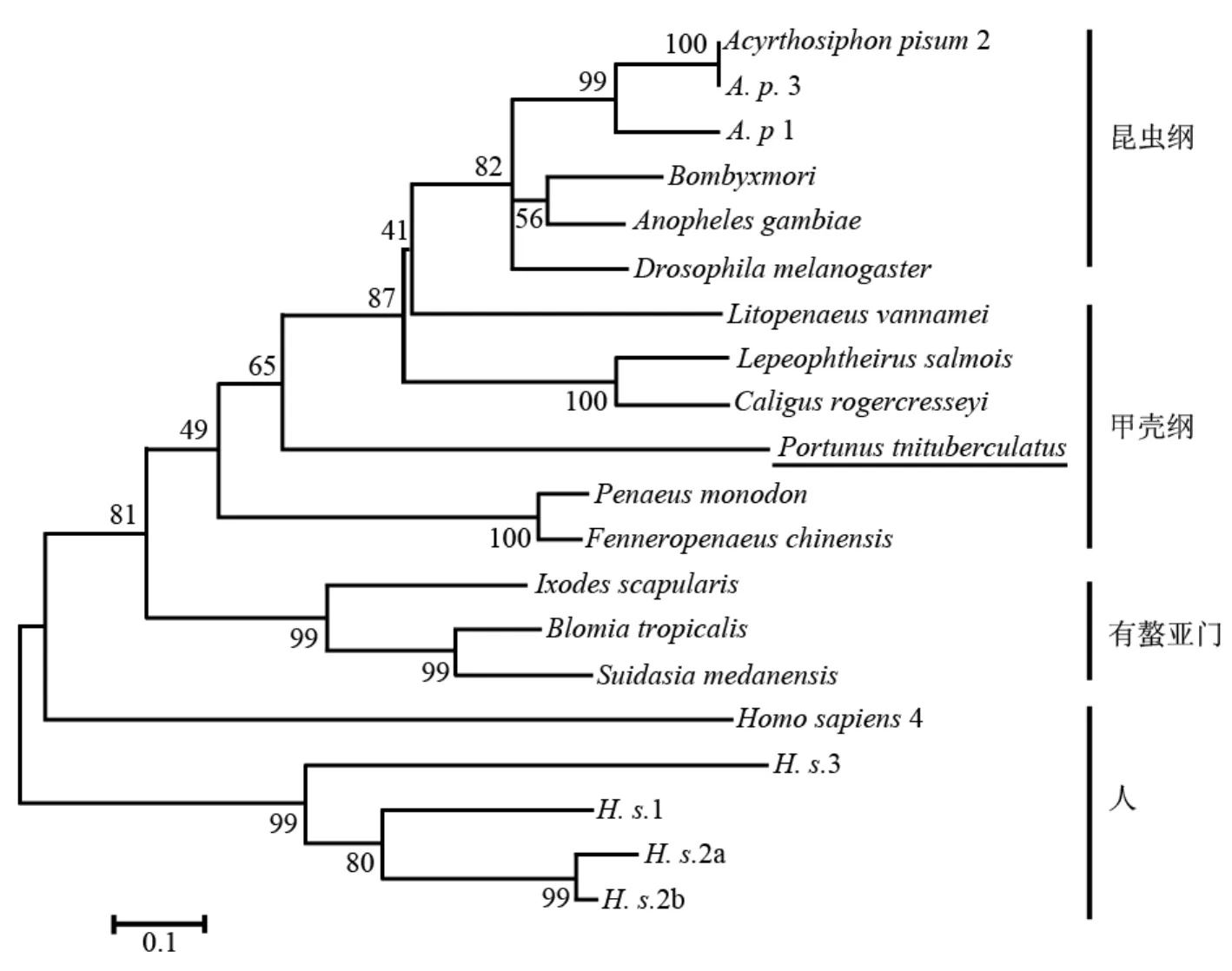
圖 3 NJ法構(gòu)建的節(jié)肢動物profilin的系統(tǒng)發(fā)生樹Fig. 3 Phylogenetic analysis of the profilins of arthropod by Neighbor-Joining method
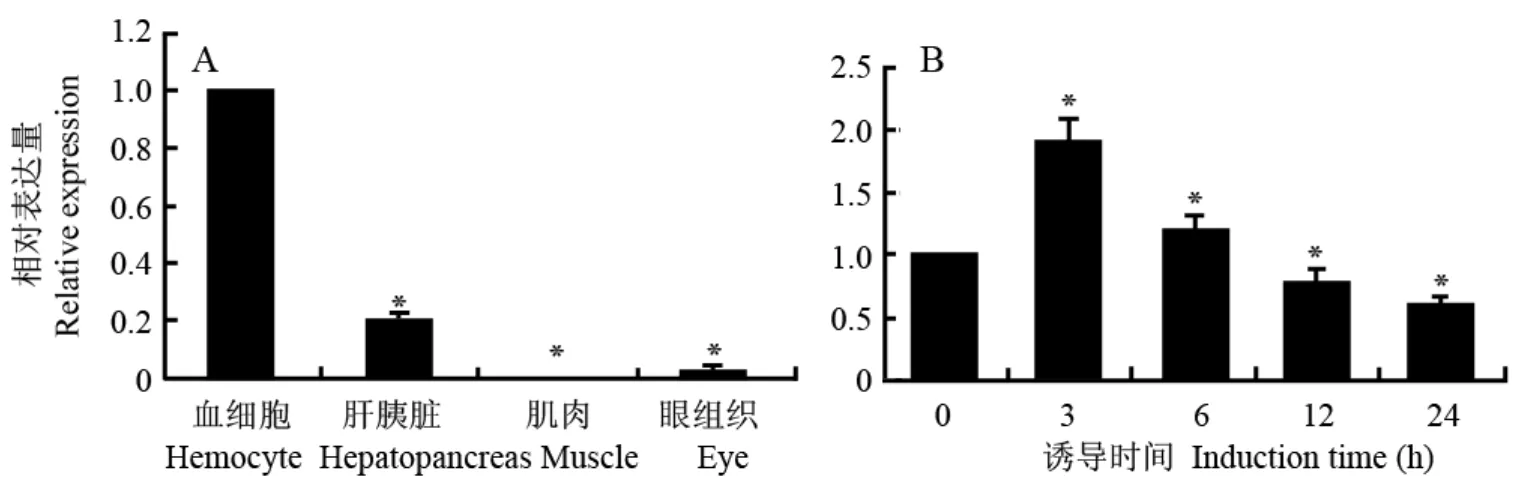
圖 4 實時熒光定量RT-PCR分析三疣梭子蟹profilin的差異表達Fig. 4 Differential expression analysis of profilin by Real-time RT-PCR
3 討 論
已有的大量研究結(jié)果表明,profilin是一種普遍存在于真核生物細胞中的小分子蛋白質(zhì),迄今為止在真核生物的各個類群,包括真菌、植物、動物等都有大量profilin被報道。同時也發(fā)現(xiàn)profilin的氨基酸序列并不保守,來源于不同物種,甚至來源于同一物種的profilin不同異構(gòu)體之間氨基酸序列同源性都可能較低(Pollard & Quirk, 1994;Obermann et al, 2005)。但profilin的二級結(jié)構(gòu)和三級結(jié)構(gòu)非常保守:4個α螺旋包圍在由7個β折疊構(gòu)成的核心的外部,形成球狀結(jié)構(gòu),在分子表面有三個配體結(jié)構(gòu)域,分別結(jié)合肌動蛋白單體(包括肌動蛋白相關(guān)蛋白,如ARP2、ARP3)、具有聚pro伸展結(jié)構(gòu)的蛋白質(zhì)(PLP配體,如Formin蛋白家族)和4,5?二磷脂酰肌醇(PIP2)等三大類配體(Mahoney et al, 1997),構(gòu)成三個配體結(jié)構(gòu)域的氨基酸序列相對保守(Huang et al, 1996),特別是N末端第三個氨基酸殘基Trp3,在所有報道的profilin中僅在人profilin IV中發(fā)生突變(Obermann et al, 2005)。本研究克隆的三疣梭子蟹profilin由125個氨基酸殘基組成,氨基酸序列比對顯示與另6種節(jié)肢動物有顎亞門動物的profilin相似性較低,氨基酸一致性只有34.9%~42.9%,但N末端相對較為保守,該區(qū)域為PLP配體結(jié)合結(jié)構(gòu)域的一部分,同時N末端第三位氨基酸殘基也為保守的Trp殘基。因此,三疣梭子蟹profilin與已報道的其他物種profilin演化規(guī)律一致,氨基酸一級結(jié)構(gòu)保守性較低,但構(gòu)成配體結(jié)合結(jié)構(gòu)域的部分區(qū)段較為保守。蛋白亞細胞定位分析結(jié)果顯示,本研究克隆的三疣梭子蟹profilin定位于細胞質(zhì),提示該profilin可能也是通過調(diào)節(jié)細胞質(zhì)肌動蛋白的動力學(xué)活性參與細胞生理過程。細胞質(zhì)中profilin在蛋白激酶的催化下,通過磷酸化與去磷酸化修飾的方式調(diào)節(jié)活性,進而影響細胞骨架的動態(tài)變化(Vemuri & Singh, 2001),在線做磷酸化位點分析顯示。三疣梭子蟹profilin共有11個可能的磷酸化位點,分別為Ser5/32/38/56/58/71/104/123、Thr84和Tyr6/125,對這些潛在磷酸位點的進一步研究,將有助于了解生理條件下profilin的動態(tài)修飾與作用機制。
熒光定量RT-PCR分析三疣梭子蟹profilin組織表達特異性結(jié)果顯示,本研究克隆的profilin主要在三疣梭子蟹免疫相關(guān)組織和器官(血細胞和肝胰臟)中表達,而在眼和肌肉中的表達量極低,分別僅為血細胞中表達量的0.028倍(P<0.01)和0.002倍(P<0.01),與中國明對蝦profilin組織表達特異性相似(Kong et al, 2009),提示甲殼類血細胞來源profilin在機體組織中的表達模式可能與甲殼類抗菌肽,如penaeidins、curstin等的表達模式類似,即主要在血細胞中以組成型表達,肌肉、眼等非免疫相關(guān)器官中檢測到的微量表達可能是組織血管中殘留血細胞造成的(Destoumieux et al, 2000; Smith et al, 2008)。三疣梭子蟹致病菌副溶血弧菌誘導(dǎo)后血細胞中profilin表達量顯著上調(diào)(P<0.01),與對蝦白斑病毒(WSSV)誘導(dǎo)后的中國明對蝦、斑節(jié)對蝦血細胞中profilin表達量變化一致(Somboonwiwat et al, 2006; Kong et al, 2009),表明本研究克隆的三疣梭子蟹profilin參與了致病微生物感染后免疫防御反應(yīng),是一個免疫相關(guān)因子。
真核生物中profilin異構(gòu)體的種類比較復(fù)雜,低等真核生物有1~3種profilin異構(gòu)體,而高等真核生物中可能更多,例如阿米巴蟲(Dictyostelium discoideum)和秀麗線蟲(Caenorhabditis elegans)各有三種profilin異構(gòu)體(Arasada et al, 2007; Polet et al, 2006),脊椎動物目前已報道的profilin有4種類型,即profilin I~IV(Polet et al, 2007),并且不同profilin異構(gòu)體有不同的組織表達特異性(Obermann et al, 2005; Polet et al, 2006)。甲殼綱已報道的profilin基因較少,在NCBI nr蛋白質(zhì)數(shù)據(jù)中僅檢索到5個profilin基因,分別來自軟甲亞綱的3種對蝦和顎足亞綱的兩種魚虱。系統(tǒng)分析顯示,三疣梭子蟹profilin與中國明對蝦、斑節(jié)對蝦profilin親緣關(guān)系較近,聚在一枝;而凡納濱對蝦profilin與兩種魚虱的profilin親緣關(guān)系更近,沒有與同屬于軟甲亞綱十足目的中國明對蝦、斑節(jié)對蝦、三疣梭子蟹profilin聚在一枝(圖3),與Kong(2009)等報道的對蝦profilin聚類關(guān)系類似。同時,本研究報道的三疣梭子蟹profilin與中國明對蝦、斑節(jié)對蝦profilin均克隆自血細胞,在外源致病微生物誘導(dǎo)下表達量均上調(diào),參與免疫防御反應(yīng),且組織表達特異性研究表明,三疣梭子蟹、中國明對蝦profilin主要在免疫相關(guān)組織血細胞和肝胰臟中表達(Somboonwiwat et al, 2006; Kong et al, 2009), 而凡納濱對蝦profilin克隆自鰓組織,在蝦白斑病毒(WSSV)誘導(dǎo)后表達量下調(diào)(Clavero-Salas et al, 2007)。因此,profilin NJ系統(tǒng)樹中甲殼綱profilin的系統(tǒng)發(fā)生關(guān)系提示,三疣梭子蟹、中國明對蝦、斑節(jié)對蝦profilin與凡納濱對蝦profilin可能分屬兩種不同類型的profilin,甲殼綱動物中應(yīng)有兩種或兩種以上profilin異構(gòu)體,并可能具有不同的組織表達特異性和生理活性。
Arasada R, Gloss A, Tunggal B, Joseph JM, Rieger D, Mondal S, Faix J, Schleicher M, Noegel AA. 2007. Profilin isoforms inDictyostelium discoideum[J].Biochim Biophys Acta, 1773(5): 631-641.
Blom N, Gammeltoft S, Brunak S. 1999. Sequence- and structure-based prediction of eukaryotic protein phosphorylation sites [J].J Mol Biol, 294(5): 1351-1362.
Bjellqvist B, Basse B, Olsen E, Celis JE. 1994. Reference points for comparisons of two-dimensional maps of proteins from different human cell types defined in a pH scale where isoelectric points correlate with polypeptide compositions [J].Electrophoresis, 15: 529-539.
Bjellqvist B, Hughes GJ, Pasquali C, Paquet N, Ravier F, Sanchez JC, Frutiger S, Hochstrasser DF. 1993. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences [J].Electrophoresis, 14: 1023-1031.
Carlsson L, Nystr?m NE, Sundkvist I, Markey F, Lindberg U. 1977. Actin polymerizability is influenced by profilin, a low molecular weight protein in non-muscle cells [J].J Biochem Mol Biol, 115: 465-483.
Chen YE, Jin S, Zhao QS. 2005. Causes and prevention ofPortunus trituberculatusdiseases in aquaculture [J].Reserv Fisher, 25(1): 78-79.[陳寅兒, 金 珊, 趙青松. 2005. 梭子蟹養(yǎng)殖中的發(fā)病原因與防治對策. 水利漁業(yè), 25(1): 78-79.]
Clavero-Salas A, Sotelo-Mundo RR, Gollas-Galván T, Hernández-López J, Peregrino-Uriarte AB, Muhlia-Almazán A, Yepiz-Plascencia G. 2007. Transcriptome analysis of gills from the white shrimpLitopenaeus vannameiinfected with white spot syndrome virus [J].Fish Shellfish Immunol, 23: 459-472.
Destoumieux D, Munoz M, Cosseau C, Rodriguez J, Bulet P, Comps M, Bachère E. 2000. Penaeidins, antimicrobial peptides with chitin-binding activity, are produced and stored in shrimp granulocytes and released after microbial challenge [J].J Cell Sci, 113(3): 461-469.
Emanuelsson O, Nielsen H, Brunak S, von Heijne G. 2000. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence [J].J Mol Biol, 300: 1005-1016.
Haugwitz M, Noegel AA, Karakesisoglou J, Schleicher M. 1994.Dictyostelium amoebaethat lack G-actinsequestering profilins show defects in F-actin content, cytokinesis, and development [J].Cell, 79: 303-314.
Huang SR, McDowell JM, Weise MJ, Meagher RB. 1996. TheArabidopsisprofilin gene family: Evidence for an ancient split between constitutive and pollen-specific profilin genes [J].Plant Physiol, 111: 115-126.
Janke J, Schlüter K, Jandrig B, Theile M, K?lble K, Arnold W, Grinstein E, Schwartz A, Estevéz-Schwarz L, Schlag PM, Jockusch BM, Scherneck S. 2000. Suppression of tumorigenicity in breast cancer cells by the microfilament protein profilin 1 [J].J Exp Med, 191(10): 1675-1686.
Jockusch B M, Murk K, Rothkegel M. 2007. The profile of profilins [J].Rev Physiol Biochem Pharmacol, 159: 131-149.
Kong HJ, Hong GE, Cho HK, Nam BH, Kim YO, Kim WJ, Lee SJ, Kim KK. 2009. Cloning of profilin (FcPFN) from the shrimpFenneropenaeus chinensis, a highly expressed protein in white spot syndrome virus (WSSV)-infected shrimp [J].J Appl Genet, 50(3): 245-250.
Kumar S, Nei M, Dudley J, Tamura K. 2008. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences [J].Brief Bioinform, 9: 299-306.
Li DP, Wang CL, Wu DH. 2008. Primary study on the resistance to emulsification disease related gene ofPortunu strituberculatu[J].J Hydroecol, 1(1): 125-128.[勵迪平, 王春琳, 吳丹華. 2008. 三疣梭子蟹抗乳化病相關(guān)基因的初步研究. 水生態(tài)學(xué)雜志, 1(1): 125-128.]
Livak KJ, Schmittgen TD. 2001. Analysis of relative rene expression data using Real-time quantitative PCR and the 2-△△Ct methods [J].Methods, 25: 402-408.
Mahoney NM, Janmey PA, Almo SC. 1997. Structure of the profilin-poly-l-proline complex involved in morphogenesis and cytoskeletal regulation [J].Nat Struct Biol, 4: 953-960.
Obermann H, Raabe I, Balvers M, Brunswig B, SchulzeW, Kirchhoff C. 2005. Novel testis-expressed profilin IV associated with acrosome biogenesis and spermatid elongation [J].Mol Hum Reprod, 11: 53-64.
Pearson AM, Baksa K, R?met M M, Protas M, McKee M, Brown D, Ezekowitz RA. 2003. Identification of cytoskeletal regulatory proteins required for efficient phagocytosis inDrosophila[J].Microbes Infect, 5: 815-824.
Polet D, Lambrechts A, Ono K, Mah A, Peelman F, Vandekerckhove J, Baillie DL, Ampe C, Ono S. 2006.Caenorhabditis elegansexpresses three functional profilins in a tissue-specific manner [J].Cell Motil Cytoskeleton, 63(1): 14-28.
Polet D, Lambrechts A, Vandepoele K, Vandekerckhove J, Ampe C. 2007. On the origin and evolution of vertebrate and viral profilins [J].FEBS Lett, 581: 211-217.
Pollard TD, Borisy GG. 2003. Cellular motility driven by assembly and disassembly of actin filaments [J].Cell, 113(4): 549.
Pollard TD, Quirk S. 1994. Profilins, ancient actin binding proteins with highly divergent primary structures [J].Soc Gen Physiol Ser, 49: 117-128.
Romero S, Didry D, Larquet E, Boisset N, Pantaloni D, Carlier MF. 2007. How ATP hydrolysis controls filament assembly from profilin-actin: Implication for formin processivity [J].J Biol Chem, 282: 8435-8445.
Smith LC, Britten RJ, Davidson EH. 1995. Lipopolysaccharide activates the sea urchin immune system [J].Dev Comp Immunol, 19: 217-224.
Smith VJ, Fernandes JMO, Kemp GD, Hauton C. 2008. Crustins: Enigmatic WAP domain-containing antibacterial proteins from crustaceans [J].Dev Comp Immunol, 32: 758-772.
S?derh?ll K, Smith VJ. 1983. Separation of the haemocyte populations ofCarcinus maenasand other marine decapods, and prophenoloxidase distribution [J].Dev Comp Immunol, 7: 229-239.
Somboonwiwat K, Supungul P, Rimphanitchayakit V, Aoki T, Hirono I, Tassanakajon A. 2006. Differentially expressed genes in hemocytes ofVibrio harveyi-challenged shrimpPenaeus monodon[J].J Biochem Mol Biol, 39: 26-36.
Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice [J].Nucleic Acids Res, 22(22): 4673-4680.
Vemuri B, Singh SS. 2001. Protein kinase C isozyme-specific phosphorylation of profilin [J].Cell Signal, 13(6): 433-439.
cDNA Cloning, Characterization and mRNA Expression of a Profilin from the Swimming CrabPortunus trituberculatus
SHEN Wang, YE Mao, SHI Ge, WANG Ri-Xin*
(Laboratory of Marine Biology Resources and Molecular Engineering, Ocean Science College, Zhejiang Ocean University, Zhoushan316004,China)
We isolated and characterized the profilin full-length cDNA from hemocytes of swimming crabPortunus trituberculatus. The profilin cDNA consists of 742 bp and the 375 bp open reading frame encodes a polypeptide of 125 amino acids, having a predicted isoelectric point of 5.87. The deduced amino acid sequence shows 42.9% amino acid sequence identity to the profilin of mosquitoAnopheles gambiae. The profilin mRNA was highly expressed in hemocytes and moderately in hepatopancreas of normal crab. The higher expression of profilin mRNA is observed in crab challenged by the pathogenic bacteriumVibrio parahaemolyticus. These results suggest a potential role for profilin in pathogen host defense mechanisms.
Portunus trituberculatus; Immunity; Gene cloning; Profilin; Gene expression
Q959.223.63; Q349; Q78
A
0254-5853-(2010)03-0261-07
10.3724/SP.J.1141.2010.03261
2009-12-22;接受日期:2010-04-27
國家科技支撐計劃(2007BAD43B08);浙江省科技廳重大項目(2007C02001);浙江省面上項目(2009C32019)
?通訊作者(Corresponding author),E-mail: wangrixin1123@126.com
申望,男,講師,E-mail: shenwangzs@163.com
book=3,ebook=178
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經(jīng)界(學(xué)術(shù)版)(2015年20期)2015-12-23 09:20:13
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
